2. СТРУКТУРА МУРАВЕЙНИКАУРАВЬИ
ОГЛАВЛЕНИЕ:
Факторы сохранения целостности семьи
Сравнимость колонн по численности населения
Колонна-муравейник в муравейнике
Облик муравьев различен. Среди нескольких тысяч видов муравьев есть и карлики размером до 2 мм, и великаны, у которых рабочие достигают 2 см. Белесые, почти прозрачные и глянцево-черные, одноцветные, двухцветные и т. д. Разнообразны форма головы и пропорция тела, скорость передвижения и способы добывания пищи. Различаются не только отдельные особи, составляющие семью, но и семьи - и по численности муравьев, и по организации общины. У одних видов это единственная самка и несколько десятков рабочих, у других - миллион рабочих и сотни самок. Чем больше население муравейника, тем сложнее организовать ее четкую жизнедеятельность. Семья может достигнуть такого уровня численности, когда существование ее как единого целого становится просто невозможным. И тогда в недрах общины начинают образовываться более мелкие внутрисемейные структурные единицы - колонны.
Чтобы сформировались колонны, необходимо несколько определенных условий. Во-первых, должна быть определенная численность колонны. Например, семья рыжих лесных муравьев не может быть слишком малой численности, так как в этом случае она оказывается нежизнеспособной. Минимум численности жизнеспособной семьи где-то около 100 тыс. Гнездо семьи из 100 тыс. муравьев имеет диаметр около 60 см и высоту 40-45 см. Такой муравейник еще слишком мал, чтобы иметь колонны (они появляются позже при численности семей 180-200 тыс. особей). Число колонн в муравейнике невелико: две, реже три (60-100 тыс. муравьев в одной колонне).
Второе необходимое условие - достаточно выраженная разобщенность колонн, прежде всего пространственная. Без этого колонны существовать не могут. Разобщение необходимо и в гнезде, и на кормовом участке. Это установлено и проверено на разных видах муравьев. В основе обособленности колонн лежит характерный для муравьев принцип вторичного деления территории, выявленный в 1956 г. польским мирмекологом Я. Добжанской. Каждый охотник действует на элементарном поисковом участке, с которым он связан длительное время. Если он и меняет участок, то это происходит только в пределах одного из секторов охраняемой территории семьи, точнее - в пределах одной дороги. Этим обусловлено постоянство сочетаний рабочих муравьев на кормовых дорогах, по которым муравьи попадают к своим поисковым участкам и колониям тлей. Сами дороги также отличаются большим постоянством, оставаясь практически неизменными 10-13 лет. Эти цифры означают лишь максимальные сроки регулярных наблюдений за одним и тем же гнездом.
Каждая дорога берет свое начало в определенной точке гнезда. Начальные точки дорог рассредоточены обычно довольно равномерно по периметру гнездового купола. Логично предположить, что муравьи, действующие на различных дорогах, придерживаются и в гнезде секторов, прилегающих к этим дорогам. Экспериментальная проверка на гнезде обыкновенного лесного муравья, произведенная нами осенью 1969 г. и летом 1970 г. в Приокско-террасном заповеднике, подтвердила такое предположение.
Фуражиры на дорогах подопытного гнезда были помечены групповыми метками. Ночью, когда все фуражиры собрались в куполе, семью усыпили эфиром. Далее был послойно разобран купол и зафиксировано местонахождение каждого меченого муравья. Оказалось, что рабочие, связанные с определенной дорогой, сосредоточиваются в прилегающей к этой дороге части гнезда. Единственным местом, где рабочие из разных колонн свободно смешиваются, оказался внутренний конус муравейника. Муравьи-строители также придерживаются своих секторов на поверхности купола. Привязанность муравьев к своим секторам гнезда оказалась очень прочной. В ходе разборки муравейника весь строительный материал купола и оказавшиеся в нем муравьи были полностью перемешаны. Когда разборка гнезда и регистрация муравьев были закончены, этот материал вместе с начавшими просыпаться муравьями собрали на прежнее место и придали ему форму, приблизительно соответствующую доопытной. И в этих условиях фуражиры и строители нашли свои секторы муравейника.
Еще нагляднее связь колонн с определенной частью гнезда у видов, сооружающих секционные гнезда, о которых говорилось выше. Там действует четкий принцип: каждой секции гнезда соответствует колонна. Пространственная разобщенность колонн сохраняется и на территории. Это вытекает уже из постоянства на территории кормовых дорог муравейника или кратеров - выходов из секций на поверхность (рис. 4, 5). Каждая колонна имеет самостоятельный выход или группу выходов на кормовой участок. На территории характер отношений колонн проявляется прежде всего во взаимодействии потоков фуражиров разных дорог.
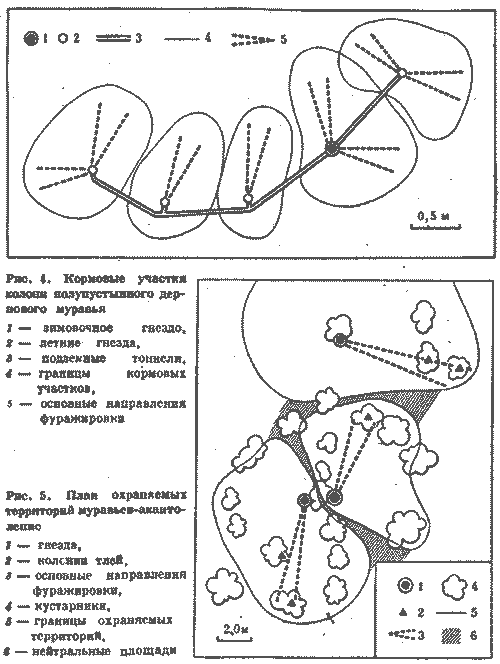
С этой целью в июне 1974 г. был поставлен специальный опыт на средней по размерам семье обыкновенного лесного муравья (диаметр купола - 0,9 м, высота гнезда - 0,6 м). Семью составляли четыре колонны примерно одинаковой мощности, о чем можно судить по сопоставимым размерам дорог (17-20 м) и интенсивности движения на них (рис. 6). После мочения на всех дорогах 2 тыс. фуражиров в зоне дороги II была установлена непреодолимая для муравьев металлическая стенка, вынуждающая поток с дороги II выйти на соседнюю дорогу I. Протяженность стенки в противоположную от дороги I сторону, к югу, превышала 15 м. Через несколько часов поток II обошел стенку с севера и вышел на полотно дороги I.
Можно было бы предполагать, что произойдет слияние двух потоков муравьев. Однако это не случилось. Войдя в вынужденное соприкосновение с потоком II, поток I отодвинулся от него к северу на 20 см. Миновав препятствие, потоки сразу же расходились, попадая в собственные русла. Такое положение сохранялось 12 дней, после чего стенка еще трижды наращивалась, пересекая каждый раз дорогу I. При этом происходили сопряженные перемещения обоих потоков к северу с сохранением установившегося между ними расстояния в 20 см. Опыт длился месяц, после чего стенка была убрана, и уже на следующий день оба потока вернулись в исходное положение.
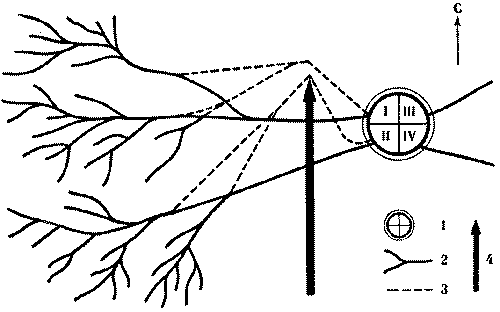
Рис. 6. Потоки фуражиров рыжего лесного муравья разных колонн «отталкиваются» при перемещении дорог.
I, II, III, IV - номера секторов и дорог гнезда; 1 - муравейник, 2 - положение дорог до и после опыта, 3 - положение дорог при максимальном смещении во время опыта, 4 - стенка
Этот опыт показал, что существует упругость дорожной сети муравейника, как бы натянутой между постоянными точками гнезда и кормового участка. Потоки фуражиров не смешиваются, они отталкиваются друг от друга. В этом выражается обособленность и целостность колонн и даже противостояние их на участке.
Обособленность колонн на территории сохраняется у рыжих лесных муравьев и в других частях кормового участка семьи, где контактируют уже не потоки, а группы муравьев. Территория каждой колонны охраняется и отделена от охраняемых территорий других колонн нейтральными пространствами - зонами, не посещаемыми муравьями (так же, как отделены охраняемые территории самостоятельных муравейников).
Кормовые участки колонн стабильны и неприкосновенны и у видов, не имеющих дорог. У обитающих в пустыне Каракум зерноядных пустынных дерновых муравьев к каждой секции примыкает свой обособленный кормовой участок. В ряде случаев он отделен от других нейтральным пространством, как и у всех зерноядных муравьев. Сбор семян производится не ежедневно, а по мере их созревания. А так как каждая колонна собирает семена самостоятельно, то обычна ситуация, когда на поверхности работают фуражиры лишь части колонн, а выходы из остальных секций закрыты. Кормовые участки закрытых секций остаются в этот день не использованными ни фуражирами из других секций данного гнезда, ни другими семьями (рис. 7).
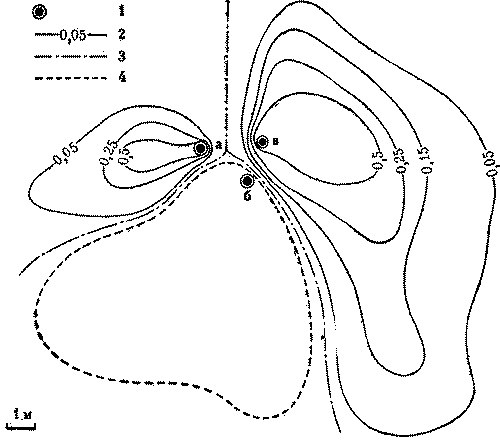
Рис.7. Использование кормового участка трехсекционного гнезда дернового пустынного муравья при работе двух секций 1 - кратеры секций а, б а в, г - изолинии динамической плотности особей на участках работающих секций а и б (особь/дм2мин), 3- границы кормовых участков секций, 4 - границы охраняемого участка неработающей секции б
Понятно, что только пространственной обособленности колонн в гнезде и на участке еще недостаточно для существования в муравейнике группировок особей. Пространственная обособленность должна сочетаться с такими важными характеристиками, как достаточно глубокая информативная изоляция и функциональная автономность колонн друг от друга. Естественно, что ни одно из перечисленных проявлений автономности колонн в рамках семьи не может быть полным, так как в этом случае семья просто перестала бы существовать как единое целое.
Информация может передаваться из колонны в колонну различными путями: посредством обмена пищей, различных сигналов (акустических, киноптических, химических, тактильных); обмена особями (личинками, куколками, взрослыми муравьями); распространения в гнезде специфического запаха, в результате чего возникает и поддерживается особый запах муравейника - гнездовой запах.
Передача различных сигналов носят прежде всего оперативный характер, в то время как обмен пищей и особями дает информацию, накапливающуюся в семье и приводящую к изменению поведения со значительной задержкой. Специфический запах семьи служит как бы эталоном при опознании своих (т. е. из своего гнезда) и чужих муравьев в гнезде и на участке.
Изучая с помощью изотопов пути распространения пищи в семьях рыжих лесных муравьев, Р.Шовен установил, что пища, добываемая муравьями на одной из дорог, не передается в другие секторы гнезда, т. е., как только в семье возникают колонны, единый пищевой поток, охватывавший ранее всех членов семьи, распадается на несколько изолированных друг от друга потоков, каждый из которых ограничен пределами одной колонны. Такая пищевая, или трофическая, изоляция колонн имеет очень важные для дальнейшего развития муравейника последствия.
Трофическая изоляция колонн и их пространственная обособленность усиливаются почти полным отсутствием передачи сведений из колонны в колонну о том, что происходит на кормовом участке. Это было проведено рядом опытов. Один из них заключался в следующем. У одиночного муравейника с помощью ловушек в течение дня отлавливали всех фуражиров с одной из дорог, интенсивность движения на подопытной дороге при этом непрерывно снижалась и через несколько часов упала до нуля в результате полной выборки всех внегнездовых рабочих из данной колонны. Казалось бы, для муравейника отлов тысяч фуражиров равнозначен гибели массы муравьев на участке одной из колонн, что означает серьезную угрозу для всей семьи.
Однако бедственное положение на одной из дорог не отразилось на скорости и интенсивности движения и поведении муравьев на остальных дорогах. Там фуражиры работали в своем обычном ритме.
Такая же картина наблюдалась и на поверхности муравейника, в контрольных секторах купола. При изъятии всех фуражиров с одной из дорог у нескольких муравейников, отличающихся по размерам и числу дорог, заметная реакция в остальных колоннах отсутствовала. Все происходящее на кормовом участке одной колонны оставалось ее частным делом и совершенно не интересовало соседей, обитающих в том же гнезде и входящих в состав одной семьи.
Абсолютен ли этот тип изоляции? Конечно, нет. Но вместе с тем настолько глубок, что мы можем проделать и более бесцеремонные действия, не вызвав тревоги у соседей. Например, надев резиновые перчатки, отлавливаем с поверхности одного из секторов купола всех находящихся там муравьев. Здесь, правда, появляется опасность, что муравьи из соседних секторов заметят резкое движение руки и устремятся навстречу надвигающейся угрозе. Однако достаточно нехитрого приспособления, чтобы этого избежать. В нижней части купола и на гнездовом валу, по границам подопытного сектора, устанавливаем невысокие стенки (5-6 см высотой). Отлов муравьев производим только между стенками. Непосредственной реакции муравьев-наблюдателей из соседних колонн на пути сборщика и боевые позы защитников гнезда из подопытной колонны уже нет. Теперь, если не делать слишком резких движений, можно отловить всех внегнездовых муравьев одной из колонн без опасений, что будет захвачено сколько-нибудь значительное число особей из соседних колонн. Муравьи подопытной колонны реагируют на исчезновение собратьев и как бы дают сигнал, когда такая выборка закончена: они быстро закрывают все гнездовые выходы. Сектор как бы вымирает. А рядом в соседних секторах все идет своим чередом: бдительные наблюдатели патрулируют поверхность купола, строители укладывают в покровный слой гнезда очередные хвоинки, непрерывным потоком идут в гнездо и вновь выходят из него потоки фуражиров. Судьба забаррикадировавшейся в своем секторе колонны никого из них не беспокоит.
Из всего изложенного можно сделать вывод, что колонны, обитающие в одном гнезде, а у многих видов имеющие даже общую крышу над головой, действуют фактически обособленно друг от друга. Но очевидно, что такое положение попросту нереально, ибо тогда непонятно, почему они все-таки обитают в одном гнезде и что их там удерживает.
Вопрос об автономности и обособленности колонн не возник бы, если бы ее состав не позволял выполнять в пределах одной колонны все основные функции, которые свойственны одинарной семье (т. е. семье, не подразделенной на колонны). В каждой колонне имеются все характерные для вида функциональные группы муравьев (и внутригнездовых и внегнездовых), выращивается молодь. Полное укомплектование колонны муравьями разных профессий и позволяет ей обособиться.
ФАКТОРЫ СОХРАНЕНИЯ ЦЕЛОСТНОСТИ СЕМЬИ
Оказывается, что в ряде случаев четкая изоляция колонн сочетается с постоянными контактами и взаимодействием в других сферах. Естественно, что эти контакты строго регламентированы. Большинство контактов имеет чисто «профессиональный» характер: строители общаются со строителями, внутригнездовые рабочие - с внутригнездовыми, фуражиры - с фуражирами. Зоны регулярного соприкосновения и взаимодействия муравьев из разных профессиональных групп пространственно разделены.
У рыжих лесных муравьев внутригнездовые рабочие контактируют во внутреннем конусе гнезда, где производится важнейшая для поддержания целостности семьи операция - регулярный обмен личинками, куколками и молодыми рабочими. Это удивительное явление чем-то напоминает меновую торговлю, но объектами обмена служат сами муравьи. По-видимому, это единственное, чем могли бы меняться колонны, ведь обмена пищей между ними нет (рис. 8).
Происходит это так. Во внутреннем конусе, являющемся настоящей инкубационной камерой муравейника (в нем поддерживается постоянная температура), обычно сосредоточен весь расплод гнезда. Каждая колонна держит молодь в своем секторе внутреннего конуса, который, однако, невелик, и расплод разных колонн оказывается рядом. И муравьи-носильщики, беспрепятственно входя в чужой сектор, забирают то личинку, то куколку и уносят их к себе. Так поступают и носильщики остальных колонн. В результате внутри муравейника идет весьма интенсивный обмен особями. Энтомолог из ФРГ Г.Кнайтп подсчитал, что в разгар сезона обмен производится каждые две минуты. Это величина достаточно большая, хотя, если учесть, что в крупном гнезде рыжих лесных муравьев одновременно может воспитываться несколько сот тысяч личинок, обмениваемая часть состава колонн относительно невелика.
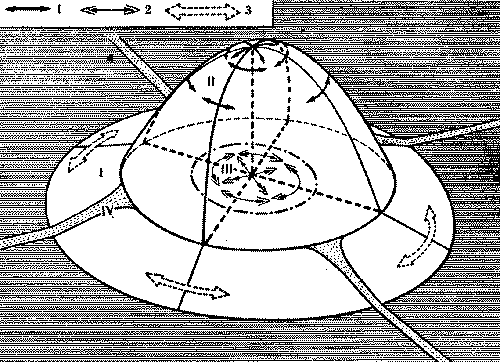
Рис. 8. Общая схема взаимодействия колонн в гнезде рыжих лесных муравьев, состоящем из четырех секций. I - гнездовой вал, II - поверхность купола. III - внутренний конус, IV - кормовая дорога. 1 - контакты муравьев-наблюдателей и строителей на поверхности купола, 2 - обмены молодью во внутреннем конусе, 3 - контакты фуражиров на гнездовом валу.
Отсутствие в гнездах многих видов внутреннего конуса не исключает внутригнездовых обменов. В секционных гнездах для этого служит система поверхностных тоннелей и камер, связывающая все входящие в состав гнезда секции. Вообще носильщики из других колонн проникают не только в «общие» помещения гнезда, но и дальше, собирая часть молоди, даже если она содержится в покровном слое или в верхних камерах секций.
Что же дают обмены особями? Они препятствуют отчуждению колонн, позволяют сохранить между ними тесные родственные отношения, исключают вражду. При обмене особями происходит скрытый обмен пищей. Все это приводит к некоторому выравниванию состава колонн как по происхождению, так и но состоянию. Сам факт устойчивого во времени существования муравейников, состоящих из нескольких колонн, говорит о том, что выработанный муравьями уровень обменов достаточен для сохранения целостности семьи.
Однако взаимодействие колонн внутри гнезда не сводится к одним обменам. Гнездо или хотя бы его общие части (внутренний конус, система поверхностных ходов и камер и т. п.) строятся муравьями из разных колонн сообща. При этом основная задача контактов сводится к обеспечению сопряжения отдельных элементов гнезда. Подобную роль играют постоянные контакты и на поверхности купола. Здесь на стыке соседних секторов взаимодействуют муравьи-наблюдатели и строители гнезда, а также фуражиры, доставляющие в муравейник материал для его сооружения. Их контакты обязательны, ибо без них муравьи не в состоянии возвести купол правильных очертаний. Это проверено экспериментально Р. Шовеном.
Ограничив с помощью нескольких стенок контакты между отдельными частями гнезда, ученый нарушил равномерность контактов строителей на поверхности купола. В результате различные части купола начинали расти неравномерно и купол утратил присущие ему округло-конические очертания. Такая картина наблюдалась даже в тех случаях, когда в каждый ограниченный стенками квадрант гнезда поступало одинаковое количество строительного материала.
Муравейник как бы накрыт контактной сетью с ячейками определенного размера. Ячейки меньше на маковке гнездового купола - здесь муравьев обычно больше и контактируют они чаще, поэтому центральная часть растет быстрее. Ближе к земле ячейки увеличиваются, скорость роста замедляется. В старых, приходящих в упадок гнездах рыжих лесных муравьев число строителей уже недостаточно для обеспечения необходимой частоты контактов. И это сразу же сказывается на форме купола. Он становится неровным, бугристым. Функция частоты контактов здесь гипертрофирована. Но, с другой стороны, представление об этакой живой форме для растущего муравейника, пожалуй, в наибольшей мере соответствует возможностям муравьев создавать впечатляющие по размерам и совершенству архитектоники целостные сооружения. Ведь общего плана муравейника нет ни у кого из его строителей.
В свое время, изучая процесс строительства галерей у термитов, известный французский энтймолог Р.Грассе выдвинул теорию стигметрии, объясняющую стереотипность элементов термитника различиями в действиях строителей на разной высоте oт пола. Такой подход поясняет и связь между размерами особей и размерами ходов и камер. Но это касается технологии сооружения отдельных элементов гнезда, соизмеримых с самими строителями. Муравейник же превосходит муравья в тысячи раз. Вот и остается муравьям разве что «взяться за руки», чтобы выстроить купол правильной формы.
Одновременно, по-видимому, решается и другая очень важная задача - соответствие размеров гнезда численности семьи. В слишком маленьком гнезде тесно, содержать очень большое - оказывается не под силу. Поэтому размеры гнезда, как правило, оптимальны для конкретной семьи. В природе задача оптимизации гнезда решается радикально: либо надстраивается купол, либо сооружаются новые секции. Уменьшение семьи влечет за собой уменьшение гнезда. Если муравьев поместить в гипсовый садок неподходящих размеров, то они либо концентрируются в части камер, если садок велик, либо «лишние» особи уходят из садка.
У самых различных видов муравьев, объединяемых лишь одним принципом общего вторичного деления территории, фуражиры составляют определенную часть семьи, в среднем около 13%. Также постоянен и процент особей, занятых уходом за самками и расплодом (25-30%). На долю строителей и муравьев-наблюдателей на поверхности купола у рыжих и тонкоголовых муравьев приходится 10-12%. На постоянстве последней величины и основывается связь между численностью семьи и размерами гнезда. Эта зависимость обнаружена и у других видов, например у строящего земляные кочки желтого земляного муравья. Даже в тех случаях, когда муравьи строят лишь небольшие временные купола для согревания личинок, сохраняется четкая связь между численностью семьи и размерами гнезда.
Имеется зона регулярных контактов и потоков фуражиров. Это гнездовой вал. Частота контактов невысока и сводится к периодическому обмену тактильными сигналами, а также к постоянному наблюдению за ситуацией в ближайшей части соседнего сектора гнездового вала. Сильное возбуждение муравьев в пограничной зоне одного из секторов привлекает туда часть фуражиров из соседней колонны. При этом отдельные фуражиры могут увлечься и попасть в чужой поток фуражиров. Такие переходы возможны только на гнездовом валу или в непосредственной близости от него. На пространственной обособленности колонн на кормовом участке все это не сказывается: потоки продолжают отталкиваться друг от друга, а сам участок остается поделенным между колоннами. Основанное на общей бдительности взаимодействие фуражиров помогает муравьям заблаговременно выявить угрозу гнезду. Надо отметить, что это всегда реакция отдельных особей или небольших групп муравьев. Чтобы активизировалась колонна в целом, источники возбуждения должны действовать непосредственно в ее гнездовом секторе. Действия колонн при этом не согласуются. Единственным местом на поверхности гнезда, воздействие на которое может вызвать общую ответную реакцию семьи, является маковка купола.
Следует отметить, что схемы взаимодействия муравьев перечисленных выше функциональных групп различны. Во внутреннем конусе каждая колонна обменивается молодью непосредственно со всеми остальными колоннами. Чтобы установить это, мы провели следующий опыт. Куколок с разноцветными метками выложили в несколько приемов на разные кормовые дороги гнезда.
Муравьи тут же начали заносить куколок в гнездо. Почти одновременно меченые куколки начали поступать в соседнее родственное гнездо, куколки с каждой новой меткой составляли определенную часть от общего числа выложенных на соответствующей дороге.
Обмен по принципу «каждая с каждой» отражает и поддерживает определенный тип взаимодействия колонн в семье муравьев. Здесь действуют многосторонние связи без какого-либо предпочтения кому-либо из партнеров.
Возможность многостороннего контакта имеется и у особей, занятых на поверхности купола. Правда, не у всех, а лишь у тех муравьев, которые функционируют на самой маковке, где смыкаются границы всех секторов. Здесь обычно максимальное число наблюдателей, так как это самое ответственное место муравейника, маковку, восстанавливают в первую очередь. Эта часть гнезда оформляется сразу же, даже если место, над которым оно должно находиться, оказалось в результате разрушения на уровне почвы.
Возможно, в какой-то мере маковка воплощает целостность муравейника.
Действительно, разделить верхнюю часть купола на две, сделать его двуглавым - значит положить начало глубокому и долгому процессу перестройки всей организации семьи. Нередко это приводит даже к разделению муравейника на два.
На остальной поверхности купола колонны контактируют и обмениваются сигнальной информацией только по эстафете. Преднамеренной передачи информации из колонны в колонну нет; муравьи воспринимают как сигналы тревоги специфические позы других особей в минуты опасности. Не имеет значения, принял ли такую позу муравей из своей или соседней колонны. Состояние возбуждения поддерживается специальными веществами - феромонами тревоги, или торибонами, выделяемыми муравьями при появлении угрозы.
Эстафетные контакты фуражиров разных колонн на гнездовом валу связаны с ролью взаимодействия особей данной функциональной группы в жизни семьи. В результате такого взаимодействия устанавливаются радиальные структуры кормового участка, а для сохранения целостности семьи контакты фуражиров значения не имеют.
Вообще гнездовой вал играет у многих видов муравьев особую роль. Здесь организационная структура семьи трансформируется в пространственную структуру кормового участка и начинается раздел сфер влияния колонн вне гнезда: определяются направления кормовых дорог или потоков фуражиров в тех случаях, когда постоянных дорог нет. Дальнейшее размежевание колонн на территории осуществляется при взаимодействии отталкивающихся друг от друга потоков фуражиров. В остальных частях кормового участка границы поддерживаются группами фуражиров, численность которых в пограничных зонах обычно выше, нежели в среднем по участку.
В итоге складывается достаточно сложная и вместе с тем рациональная система постоянного взаимодействия колонн в гнезде и на территории, схематично изображенная на рис. 8. И хотя колонны обособлены и во многом развиваются почти независимо друг от друга, они все-таки составляют единое целое - семью.
Подытоживая изложенное в этом разделе, следует назвать два фактора, наиболее важных для сохранения семьи муравьев: общее гнездо и регулярные обмены особями Гнездо важно не только как жилище, общая крыша. В таком гнезде присутствует и постоянно выравнивается единый запах семьи. Ведь именно по запаху муравьи отличают своих от чужих. Пока муравейники имеют одинаковые или близкие запахи, воины между ними не будет. Встретившись на куполе или вдали от него рабочие муравьи первым делом обмениваются легкими ударами антенн. В булавах антенн находятся хеморецепторы, с помощью которых муравей и определяет запах различных предметов, в том числе и запах муравьев. Свой будет отпущен или привлечен к какой-нибудь деятельности. Чужому не поздоровится; его либо разорвут на части, либо изгонят.
Обмены как средство сохранения лояльных отношений колонн дополняют общность гнезда и общность гнездового запаха.
Однако обмены оказываются крайне необходимыми при выполнении еще одного определяющего условия устойчивости семьи: сравнимости колонн по численности.
СРАВНИМОСТЬ КОЛОНН ПО ЧИСЛЕННОСТИ НАСЕЛЕНИЯ
Сколько-нибудь выраженной иерархии колонн в пределах семьи нет. В секционных гнездах это обусловлено тем, что все секции единого гнезда равны между собой. Одинаковы по численности и колонны, населяющие секции. Это показано на разных видах, относящихся к различным родам и подсемействам.
Численность населения одной секции стабильна для каждого вида в конкретных условиях обитания. А поскольку отношения группировок муравьев одного вида отражают в конечном счете соотношение численности этих группировок, то основа для доминирования какой-либо из колонн в семьях, обитающих в секционных гнездах, отсутствует.
У стабилизированной семьи рыжих лесных муравьев все обстоит сложнее, так как с сооружением купола численность муравьев перестает зависеть от емкости подземной части гнезда. Купол можно надстроить, увеличив его диаметр и высоту. Увеличился объем купола - возросли возможности гнезда вместить дополнительное количество особей. Поэтому какой-то определенной численности колонны здесь нет.
В гнезде одного размера может быть разное число колонн. Так, у северного лесного муравья в гнезде с диаметром купола 1 м оказывается и две, и пять колонн. Естественно, что в первом случае численность каждой колонны выше. Но внутри семьи колонны, как правило, соизмеримы. И чем старше муравейник, чем меньше вокруг других гнезд, так или иначе влияющих на его рост, тем ровнее длина дорог, интенсивность движения на них и общая численность колонн. В идеальной ситуации, когда одиночный муравейник расположен в равномерном по составу, возрасту и продуктивности насаждении, кормовой участок приобретает форму круга.
Такие семьи обладают наибольшей устойчивостью во времени, и только серьезное вмешательство извне может вывести их из стабилизированного состояния. В этом случае чем больше колонн в муравейнике, тем меньше вероятность последующих серьезных изменений в структуре семьи. И все же такие изменения происходят. Гнездо растет, и у него появляются новые дороги. На некотором расстоянии от старого муравейника возникают новые муравейники, которые тоже начинают расти. Наступает время, и здесь образуются колонны.
Что же приводит к этим изменениям? Ведь очень часто новые колонны возникают задолго до того момента, когда уже имеющиеся достигли верхнего предела численности. Такой предел еще не известен, однако в совершенно одинаковых условиях по соседству существуют муравейники с разной численностью колонн. Например, у каждого по три колонны, но колонны первого гнезда насчитывают по 50-60 тыс. особей, второго - 90- 100 тыс., а третьего -150-200 тыс. особей. Можно было бы ожидать, что новая колонна возникнет прежде всего в последнем, наиболее мощном муравейнике. Однако в действительности очень часто дело обстоит иначе. Следовательно, абсолютная численность еще не определяет ситуации.
Чтобы выяснить, в чем причина, был проведен следующий опыт. В 1969 г. мы подобрали шесть близких по размерам муравейников обыкновенного лесного муравья, имевших по три равных между собой дороги. Территория каждого подопытного муравейника была тщательно заквартирована до начала опыта. Все шесть муравейников были одиночными. Четырем из них на одну из дорог в течение июля-августа выкладывали в больших количествах куколок того же вида, взятых из гнезд, не участвовавших в опыте. В несколько приемов в каждом случае на подопытную дорогу было выложено по 100 тыс. куколок.
Вышедшие из куколок рабочие муравьи влились в состав соответствующей колонны, которая таким образом резко усилилась. Два других муравейника в это же время также были усилены куколками, но одновременно все колонны усиливались на 90-100 тыс. особей каждая.
Реакция муравейников в первом и втором вариантах оказалась различной. Равномерное усиление всех колонн привело к росту гнезд и также равномерному увеличению дорожной сети каждой колонны. Усиление одной из колонн нарушило имевшееся до опыта равновесие. Кормовой участок такой колонны резко расширился, дорога удлинилась и появились новые ответвления. Контрольные дороги не изменились. Наконец, одна из усиленных колонн разделилась и выделила дочернюю семью. Отводок был сооружен в сентябре того же года, через месяц после усиления колонны куколками.
Из приведенного опыта следует несколько выводов. Обмены, которые производят муравьи внутри гнезда, не обеспечивают выравнивания численности колонн в случае резкого усиления одной из них. Если одна из колонн вследствие каких-либо причин начинает расти быстрее остальных, различия в численности накапливаются. В определенный момент подобная несоразмерность может послужить причиной выделения отводка наиболее сильной колонной.
Интересно, что в различных экспериментах и при естественном ходе событий дочернюю семью всегда выделяет самая мощная колонна, резко выделяющаяся по численности особей. Есть все основания расценивать образование отводка как средство сохранения структуры семьи: проблема внутрисемейной регуляции решается путем расселения и создания новой семьи.
Образование отводка - один из путей выравнивания колонн по численности населения, наиболее радикальный, но не единственный. Второй, более обычный путь - формирование новой колонны. Здесь тоже обязательным условием является нарушение численного равновесия колонн.
Образование новой колонны теснее связано со структурой кормового участка, с дорожной сетью муравейника. Возникновение колонны у рыжих лесных муравьев невозможно без появления новой дороги. Число разветвлений у той или иной дороги роли не играет. Важно не число ответвлений на территории, а число потоков, выходящих из гнездового купола. Поэтому процесс разделения колонны начинается всегда на кормовом участке, схематично он изображен на рис. 9. Из схемы видно, что разделение в большей степени зависит от возможности радикального раздвоения исходного кормового участка колонны. Чем шире занимаемый участком сектор, тем проще использующей его колонне разделиться на две. Число колонн и связанных с ними дорог у рыжих лесных муравьев весьма постоянно. Еще в 30-х годах появились работы немецкого энтомолога Р. Штегера, проследившего топографическое постоянство кормовых дорог в течение 13 лет. Имеются подобные данные и у других авторов, работавших с муравьями разных родов. Смещения отдельных ответвлений дорог обычно не связаны с изменением в составе колонн. А иногда разделение колонны на две почти не отражается на периферийных отрезках дорог (именно этот вариант показан на рис. 9, в).
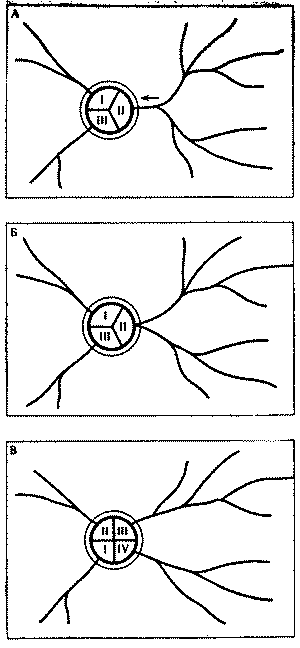
Рис. 9. Процесс образования новой колонны при раздвоении дороги
А -
наметившееся разделение наиболее сильной и
разветвленной дороги (стрелка показывает
направление перемещения развилки);
Б - положение накануне разделения колонны
на две: развилка переместилась к основанию
купола;
В - колонна разделилась, ответвления стали
самостоятельными колоннами
I, II, III, IV - номера колонн (дорог, секторов)
Увеличение колонн - процесс, направленный как бы внутрь муравейника, ведущий к умножению связей в семье, укрепляющий ее как единое образование. А это означает, что данный процесс в какой-то мере противодействует почкованию - выделению дочерних муравейников. Действительно, чем больше колонн, тем прочнее каждая из них связана с семьей, тем труднее одной из них достичь условий, необходимых для выделения отводков.
Наибольшее значение имеют при этом два показателя: размер семьи, тесно связанный с размерами муравейника и кормового участка, и число колонн. В результате взаимодействия двух тенденций вырабатывается оптимальная зона образования отводков. Причем эта зона даже не видоспецифична вообще, а отражает специфику вида лишь в определенных условиях. Так, в подмосковном ельнике у обыкновенного лесного муравья максимум образованных отводков падает на муравейники с диаметром 1,2-1,3 м, состоящие из четырех колонн. А в сосняках у того же вида диаметр наиболее плодовитого гнезда-1-1,1 м при трех колоннах.
Процессы разделения колонн и почкования противодействуют друг другу и имеют разные последствия для роста гнезда. Увеличение числа колонн - это путь к развитию крупных муравейников, появлению «сверхгнезд». Муравейники-гиганты почти всегда одиночные и всегда включают много колонн. Максимальное число колонн в средней полосе отмечено нами в Приокско-террасном заповеднике; семья малого лесного муравья имела 12 дорог. Охраняемая ею территория занимала более 1,5 га. Почкование, наоборот, сдерживает рост муравейника. Причем сдерживание происходит за счет самой мощной, т. е. наиболее быстро растущей колонны, что особенно важно. В итоге семья удерживается в пределах, соответствующих состоянию интенсивного выделения отводков.
Попробуем охарактеризовать этот размерный интервал с позиции оптимизации размеров семьи. Представим себе требования, которым должен соответствовать такой муравейник.
В общих чертах оптимальной организацией любого биологического объекта можно считать такую организацию, которая обеспечивает устойчивую плотность поселения на максимальном для данных условий обитания уровне. Это почти в равной мере применимо к различным уровням организации. Для муравьев мы вправе использовать такой подход по отношению к отдельной семье, комплексу муравейников одного вида и даже к обитающим на одной территории поселениям муравьев разных видов, или, как их назвал в 1975 г. московский мирмеколог А.В.Демченко, к многовидовым ассоциациям муравейников.
Для отдельного муравейника основным показателем оптимальности организации будет стабильность размеров семьи и гнезда во времени. Важны также вероятность гибели муравейника определенного размера, его возможности к дальнейшему росту и образованию отводков. Необходимо выбрать отрезок времени, соответствующий нашей задаче. Год для этой цели слишком малый срок: условия разных лет могут резко отличаться друг от друга, а это неизбежно сказывается на состоянии каждой семьи. Но пятилетнего периода уже достаточно, чтобы выявить общие тенденции развития муравейников. Вот что дал анализ пятилетних данных по 112 семьям северного лесного муравья, обитающего в ельнике-кисличнике в районе Солнечногорска Московской области.
Совсем маленькие муравейники (диаметр до 40 см), как правило, обречены. Если в течение двух лет такой муравейник не вырастет в более крупный, он погибнет. Погибает 83% подобных гнезд. Муравейники этого размерного класса не обладают устойчивостью. Среди муравейников покрупнее (диаметр 45-60 см) доля погибших меньше (24% за пять лет), но и здесь семья всегда перед альтернативой: либо вырасти, либо погибнуть. Лишь 4% семей сохраняется в исходном классе в течение пяти лет. Зато большинство муравейников (72%) вырастают, причем некоторые за это время достигают метра в диаметре.
Далее вероятность погибнуть уменьшается, но скорость роста начинает снижаться. Одновременно появляется и обратная тенденция: муравейники, продержавшись некоторое время в более высоком классе, вновь попадают в разряд более мелких. Это особенно заметно на примере крупных гнезд диаметром 1,25 м и более. Тенденции к росту резко падают с увеличением контрольного периода с одного до пяти лет. Такой муравейник может выйти в следующий класс на один-три года, но в течение пяти лет ни один из них не удерживается там и снова уменьшается в размерах, нередко даже ниже исходного уровня. Среди гнезд диаметром более 1,4 м в своем размерном классе удерживаются только из муравейников, а остальные - начинают деградировать.
Единственной устойчивой размерной группой оказались муравейники с диаметром купола 1,05-1,2 м. Большинство муравейников этого размера сохраняются в классе независимо от срока наблюдения. У них еще сильна тенденция к росту: почти 1/3 гнезд за пятилетие переходит в следующий размерный класс. Это значит, что муравейники такого размера, обретая устойчивость, сохраняют активность. Вероятность гибели муравейников данного класса за пятилетний период равна нулю. И лишь 7% гнезд уменьшается в размерах.
Но, пожалуй, самое интересное заключается в том, что муравейники именно этого диаметра образуют основную часть естественных отводков в комплексе. Следовательно, есть все основания рассматривать образование отводков как механизм поддержания численности семьи на определенном, оптимальном для данного вида в данных условиях уровне.
Что же это за уровень? Расчеты показали, что в гнезде северного лесного муравья диаметром 112,5 см (середина класса) обитает около 870 тыс. муравьев. Эти муравьи организованы, как правило, в четыре колонны (среднее число колонн - 4,11). -
Знакомясь с двумя механизмами стабилизации муравейников, мы не должны упускать из виду одну важную общую черту. Действие этих механизмов непостоянно. Они срабатывают периодически, как ответная реакция семьи на накопившиеся количественные диспропорции между составляющими ее колоннами. И образование отводка, и формирование новой колонны - специфические аварийные меры сохранения семьи. И хотя фактологическая сторона процессов достаточно прояснилась, один из наиболее интригующих, узловых моментов остается неясным: какие реальные силы заставляют самую могучую колонну в семье выделять отводок? Почему именно так решается этот важный для муравейника вопрос? Ведь заглядывая немного вперед, можно сказать, что образование отводков далеко не всегда выгодно материнскому муравейнику. Казалось бы, усилившаяся колонна могла бы потеснить соседей по гнезду или просто расширить свой сектор.
К сожалению, ответов на эти вопросы еще нет. Очевидно только, что существенного передела секторов гнезда в обычных условиях не происходит, так же никогда не наблюдаются внутригнездовые муравьиные войны.
В природе мы можем застать самые различные этапы оптимизации муравейника. Этот процесс складывается из двух процессов: оптимизации размеров семьи и оптимизации ее организации. Они взаимосвязаны, дополняют друг друга, но не совпадают. Синхронности здесь тоже нет, потому что первый процесс почти непрерывен, а второй - четко дискретен. Поэтому первый процесс обычно идет быстрее. Численность колонн «догоняет» его скачками (и даже может опередить).
При этом увеличение числа колонн связано с формированием наиболее рациональной формы кормового участка - округлой.
Формированию кругового кормового участка могут препятствовать различные факторы. Такой участок маловероятен при неравноценности кормовых угодий вокруг муравейника. Распределение растений с колониями насекомых, выделяющих падь, также имеет значение. Особенно сильно сказывается размещение соседних муравейников. Конечно, не всяких, а того же или конкурирующих видов. Установив в 1932 г. наличие у муравьев охраняемой территории, английский эколог Ч. Элтон положил начало целому направлению в изучении жизни муравьев - первых насекомых, у которых данный вопрос был разносторонне изучен.
Конкуренция сводится к борьбе за территорию, реже - за места поселения. Кормовые участки расположенных поблизости муравейников вытянуты в противоположные стороны либо параллельными полосами. Нередко форма их причудлива: кормовые дороги с прилегающими полосками угодий как бы пробираются среди чужих земель. В таких условиях выровнять колонны путем увеличения их числа нельзя. И тогда выручают отводки. Вместе же оба процесса дают большинству семей достаточно богатые возможности для достижения устойчивого состояния.
КОЛОННА-МУРАВЕЙНИК В МУРАВЕЙНИКЕ
Достигая определенных размеров, семья муравьев начинает дробиться на части, каждая из которых вполне автономна и в состоянии полностью или почти полностью выполнять все необходимые для полноценного существования функции. Под одной крышей сосуществуют несколько вполне сформировавшихся муравейников? - Именно так и обстоит дело. Еще раз перечислим основные характеристики, указывающие на самостоятельность колонн. Сюда относятся пространственная и пищевая обособленность в гнезде и на кормовом участке, устойчивость состава, отсутствие передачи из колонны в колонну оперативной информации о состоянии и делах на территории, фактически автономное развитие.
У полигинных видов каждая колонна в состоянии перейти в самостоятельное гнездо. Это довольно часто происходит при сильном разрушении муравейников (например, кабанами). На месте разрушенного гнезда, на сохранившемся гнездовом валу или неподалеку от него, возникают несколько муравейников. Число таких гнезд обычно равно числу колонн в исходной семье. Иногда из семьи уходит только одна колонна, нередко часть колонн объединяется в одном из вторичных муравейников, а часть - в другом. При таких внутренних разделах соблюдается в общем-то лишь одно правило: гнезда меняются, семьи меняются, а колонны остаются, сохраняя свой кормовой участок и дорогу.
Но силы, ранее удерживавшие их в одном гнезде, сохраняются. По крайней мере первое время. Впрочем, понятие «первое время» здесь относительно, оно может растянуться и на несколько лет. Бывают и быстротечные варианты, когда уже через две-три недели все отношения между бывшими сородичами прекращаются. Это случается чаще при обосновании каждой колонной своего гнезда. Связи между групповыми вторичными муравейниками гораздо устойчивее.
Дальнейшая судьба вторичных муравейников различна. Колонны, прервавшие связи, становятся самостоятельными семьями и далее развиваются как одиночные муравейники. Если связи между гнездами устойчивы, события принимают почти фатальный характер. Вторичные гнезда исходно равны или почти равны друг другу, так как равны основавшие их колонны. Некоторое время, не более одного-двух лет, это равенство сохраняется. Между муравейниками существует оживленная дорога с интенсивным взаимным обменом особями. Обмен всегда равный - один к одному. Семьи продолжают придерживаться принципа, действовавшего, когда они еще были колоннами одного муравейника.
Но колонны развиваются с разной скоростью. И семьи тоже. Один муравейник растет немного быстрее, другой - медленнее или вообще не растет. Спустя некоторое время различия между ними становятся настолько существенными, что меньший становится чем-то вроде отводка у более крупного. И тогда более крупный поглощает, присоединяет его. Население маленького гнезда переселяется при этом в большое. Но опять же колонны остаются. Этапы такого воссоединения в одном гнезде схематично изображены на рис. 10. Отметим, что все это возможно только у относительно небольших одинарных муравейников. Если муравейники стали взрослыми и в них произошло разделение колонн, подобная ситуация уже исключена.
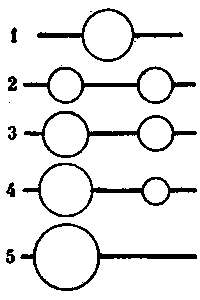
Рис.10. Воссоединение колонн, перешедших в отдельные гнезда после разрушения исходного муравейника
1 -
исходный муравейник с двумя колоннами,
2 - равные по размерам гнезда колонн
непосредственно после обособления,
3 - нарушение равенства колонн в
результате их неравномерного роста,
4 - различия в силе колонн достигают
критической величины,
5 - колонны объединились в более крупном
муравейнике, вслед за этим часто следует
деление усилившейся- колонны.
На первый взгляд объединение, в общем гнезде при таких обстоятельствах противоречит изложенному выше принципу выравнивания колонн путем выделения отводка. Там выделение, а здесь та же неравноценность колонн ведет к объединению. Однако противоречия нет. Отводок должна выделить наиболее сильная колонна, входящая в муравейник. Но если колонна в муравейнике одна, ей не с кем сравнивать свою мощь. Система из двух одноколонных муравейников после усиления одного из них начинает несколько напоминать материнский муравейник и отводок. Но и это лишь внешнее подобие, так как отводок может возникнуть только у муравейника с несколькими колоннами, т. е. ситуация искусственная и именно поэтому неустойчивая.
И все-таки система существует один-два года?! В том-то и дело, что неустойчивой система становится лишь после нарушения равновесия разбежавшихся по отдельным гнездам колонн. Когда различия в размерах семей становятся кратными, поглощение меньшей происходит быстро, как правило во время весеннего или осеннего массового обмена между гнездами. Такие обмены нередко интенсивнее летних. Поэтому они и являются обычно как бы поворотным пунктом в жизни муравейников.
В объединении муравейников проявляется очень сильная тенденция муравьев к концентрации. Об этом мы уже говорили в первой главе. Она даже доминирует при отсутствии взрослых муравейников, например, после искусственного переселения отводков. Аналогичная ситуация складывается и в результате распадения семьи на несколько мелких. Концентрация муравьев протекает одинаково в обоих случаях. Следствием объединения семей-недоростков в одном гнезде оказывается последующий быстрый рост этого муравейника. Причин тут, видимо, немало: улучшение температурного режима в гнезде (в большом гнезде он успешнее выдерживается), повышение конкурентоспособности семьи и, что, наверное, самое главное, восстановление естественной структуры семьи. По-видимому, мы не ошибемся, если назовем подразделение большой семьи ряда видов на колонны обязательным условием их нормального существования и развития. Семья, состоящая из одной колонны, - всего лишь начальный этап жизни муравейников многих видов.
А что же происходит в тех случаях, когда «разбежавшиеся» колонны неопределенно долго остаются равными по численности? Этот вариант тоже ведь не исключен. Встречается и такое: два муравейника растут рядом, связанные обменной дорогой. Со временем в каждом из них возникают новые колонны, иногда дело доходит и до образования отводков. А давняя дорога продолжает существовать. Впрочем, не бесконечно долго. В конце концов муравейники-близнецы обособляются, и дорога между ними зарастает травой.
Таким образом, каждая колонна настолько индивидуальна и автономна, что всегда может начать самостоятельное существование. Но все-таки одноколонный муравейник, по-видимому, нерационален. Именно поэтому колонны образуются уже в небольших муравейниках. Общая численность населения такого гнезда находится на уровне 200 тыс., т. е. даже меньше численности одной колонны процветающего муравейника.
Имеется масса видов с небольшими семьями, которые никогда не достигают уровня, необходимого для формирования колонн. Сюда относятся муравьи с наиболее простой организацией и зачастую невыраженной структурой семьи. Для некоторых видов сама возможность образования внутрисемейных группировок особей открыта, но реализуется только в исключительных случаях. Есть и такие виды, у которых достижение семьей определенной (пороговой) численности служит сигналом к разделению ее пополам (гесмозису). Примерно так, как это происходит у медоносной пчелы. Только события, естественно, разворачиваются в гнезде или на поверхности почвы. Гесмозис у муравьев рода афеногастер описан французским энтомологом А.Леду в 1973 г. и рода мономориум - индийскими мирмекологами супругами Сеансами в 1971 г.
При делении старая самка вместе с частью рабочих покидает свой муравейник и основывает новый. В оставшемся гнезде из отложенных старой самкой яиц выкармливаются молодые самки, одна из которых после оплодотворения начинает яйцекладку. Эти виды монотонные, поэтому гесмозис связан с обеспечением самками обеих частей делящейся семьи. Этим обусловлено столь значительное сходство гесмозиса у данных видов муравьев и у пчел. Но гесмозис происходит и у некоторых полигинных муравьев, например у повсеместно распространенных у нас мирмика. Деление семьи мирмика, прослеженное А. В. Демченко, происходило по упрощенной схеме: самки, так же как и рабочие, делятся на две группы, после чего одна из группировок покидает старое гнездо и переходит в новое, сооружение которого начинается заблаговременно частью рабочих, муравьев.
Здесь, как и в гнездах формика, длительный процесс наращивания численности ведет к вызреванию в недрах процветающей семьи более мелких и соизмеримых с видовым оптимумом объединений индивидов. Различия в том, что семья не усложняется структурно, а распадается на две. Это происходит, очевидно, как следствие относительно слабых внутрисемейных связей. Отсутствует та могучая центростремительная сила, которая, как мы видели на примере рыжих лесных муравьев, не только доминирует над автономными силами колонн внутри гнезда, но и способна удержать колонны в рамках общей системы даже при его распадении. Вероятно, развитие механизмов, обеспечивающих сохранение целостности семьи при делении семьи на колонны, является тем эволюционным достижением, который по своему значению для дальнейшей судьбы муравьев намного превосходит даже сам принцип деления.
К этому вопросу мы еще вернемся, а пока лишь отметим, что в любом варианте деление общины происходит по дихотомическому принципу, что образующиеся при этом новые семьи или колонны - организационно завершенные и жизнеспособные единицы, способные к самостоятельному существованию. И, наконец, первая задача, которую решает в такой ситуации семья, - это оптимизация своей численности и организационной структуры. Образование новых муравейников в стратегию муравейника не входит, а является следствием решения первой задачи.
Необходимо отметить, что образование колонн резко повышает жизнеспособность каждого муравейника и комплексов муравейников. Это особенно важно для видов, сооружающих наземные купола из легких строительных материалов. Такие гнезда не защищают их обитателей от крупных млекопитающих: медведей, барсуков, кабанов, - а, наоборот, привлекают их. В этих условиях рассредоточение колонн по отдельным гнездам - единственное средство пассивной защиты, которое могут противопоставить муравьи. Небольшие муравейники звери почти не трогают. В результате одним из следствий массовых разорении муравейников кабанами оказывается увеличение общего числа гнезд при уменьшении их средних размеров.
В арсенале ряда видов имеются и некоторые другие средства выхода из критической ситуации, когда семья разрастается до таких размеров, что нормальное существование ее как единой системы индивидов становится невозможным. Речь об этом пойдет в последнем разделе данной главы.
Итак, можно охарактеризовать семью муравьев как систему, состоящую из нескольких идентичных по составу и сравнимых по численности особей подсистем - колонн, которые имеют постоянный состав, занимают определенные части гнезда. Эти части (секции, секторы) подобны друг другу. Кормовой участок, используемый каждой колонной, также постоянен.
Колонны сравнимы по численности, и их отношения строятся не на иерархическом принципе, а на многостороннем равнодействии всех входящих в систему подсистем. Возможность проявления иерархических отношений исключается, так как вслед за непомерным усилением одной из колонн неотвратимо следует либо разделение этой колонны на две, либо выделение части ее населения в отводок. Муравейник действует как целостная система, хотя какие-либо специальные образования, координирующие рост и жизнедеятельность колонны или управляющие ими как подсистемами целого, отсутствуют.
Перечисленные свойства присущи особому координационному типу систем, представляющему собой довольно низкий уровень интеграции. На этом уровне в наименьшей степени проявляется зависимость подсистем друг от друга и от системы в целом, что мы и видим на примере муравьев. Общесемейные реакции формируются путем прямого сопоставления колонн между собой и каждой из них со всеми остальными.
Однако имеются и некоторые серьезные отличия системных свойств муравейника от традиционных координационных систем, например популяции. Из них стоит выделить три.
Во-первых, это ограниченное число подсистем, входящих в муравейник. Обычно 2-5, максимально 12, в то время как в любой благополучной популяции тысячи и миллионы индивидов. Немногочисленность подсистем повышает удельный вес каждой из них, делает значимым для состояния семьи любое изменение их количества. Одновременно функциональные связи между всеми колоннами становятся обязательными и регулярными. Прекращение таких связей означает выпадение колонны из семьи, чего в нормальных условиях не бывает. Все это приводит к значительно большей организованности муравейника как одного из вариантов координационных систем.
Во-вторых, колонны объединяются не только функциональными связями, но и общим местом обитания - гнездом, которое является основным выражением интеграции их деятельности. В общем гнезде регулярные связи всех колонн обретают важное дополнительное свойство: они осуществляются по строго определенным путям. И если в муравейнике нет координирующего функционального образования, какой-нибудь колонны-координатора, то там имеется определенная зона (внутренний конус или группа поверхностных ходов и камер), где сосредоточены основные пути координации. Это дополняет вытекающие из первого положения системные возможности семьи муравьев, еще более укрепляя ее организационную целостность, делая ее функционально более эффективной.
В-третьих, система контролирует изменение числа колонн уже на начальном этапе, регулируя их численность и размеры. Благодаря этому появилась возможность сопряженного устойчивого роста подсистем и семьи как объединяющего их целого. Почти стихийный рост колонн преобразуется в результате их взаимодействия в зарегулированный рост семьи. На определенном уровне численности появляется стабильность. Ее обеспечивает высокая степень организации муравьиной общины. Без четкой организации существование муравьев в таких количествах было бы просто невозможно.
Для более полной характеристики муравейника как системы требуется привлечение данных о системных свойствах самих колонн, т. е. об их поведенческой структуре. Этот чрезвычайно емкий и очень интересный вопрос не входит в нашу задачу. Однако стоит упомянуть, что поведенческая структура колонн достаточно сложна и, как правило, более видоспецифична. Все моменты, связанные с разнообразием каст и размерных групп муравьев, со сменой профессий и взаимодействием профессиональных групп между собой, добычей пищи и ритмикой общей активности, с особенностями поведения муравьев в гнезде и на участке, регулируются в рамках отдельной колонны.
Об организационной структуре муравейников известно еще очень мало. Многие вопросы требуют тщательной разработки, другие - едва лишь затронуты исследователями. Ограничен и круг видов, используемых в качестве модельных для этих целей. Но все-таки уже сейчас можно, хотя бы в первом приближении, представить себе, сколь сложна и совершенна организация семьи муравьев, насколько своеобразен их мир, преодолевший не одно, казалось бы, непреодолимое препятствие - мир, где собранные по крупинкам достижения индивидов синтезированы в строгие и действенные формы организации.
Для рациональной организации семьи и при перенаселенности гнезда у муравьев наблюдается не только образование колонн. В арсенале ряда видов имеются некоторые другие возможности выхода из критической ситуации. В настоящее время мы имеем сведения еще о двух типах внутрисемейной организации - это поликалия и диффузные гнезда.
Поликалия - обитание одной семьи в нескольких взаимосвязанных гнездах - очень широко распространена у муравьев, в первую очередь среди видов, в семьях которых имеется только одна самка. Помимо основного, самого крупного гнезда муравьи сооружают вспомогательные, находящиеся на некотором расстоянии от основного и выполняющие различные функции. Эти гнезда гораздо меньше по размерам и могут существовать различное время: несколько дней, в течение одного сезона, ряда лет или всего срока существования семьи.
Изучение поликалии начато еще в прошлом веке выдающимся французским мирмекологом А.Форелем. С тех пор описаны вспомогательные гнезда, различающиеся по своим размерам, сроку существования и назначению. Небольшие временные гнезда муравьи строят при переселении на новое место, пряча там расплод, самок и внутригнездовых рабочих. Во второй половине лета большинство муравьев, обитающих в умеренном поясе, переключаются на сбор пади корневых тлей. При этом для каждой колонии тлей строится небольшое убежище, состоящее из нескольких камер различных размеров. В степи корневые тли - основной источник пади для муравьев. Здесь убежища тлей почти постоянны. В каждом убежище находится небольшой постоянный гарнизон рабочих, поддерживающих гнездо в порядке и охраняющих его. Убежища для тлей существуют около двух месяцев. Пахучие муравьи-древоточцы накрывают колонии тлей картонным чехлом. Постоянно укрывают колонии тлей в галереях рабочие желтого пахучего муравья.
Чтобы пережидать непогоду, муравьи сооружают микрогнезда, или, как их называл А.Форель, «станции» и «павильоны». Такие укрытия часто находятся в стороне от дорог. Еще одним типом вспомогательных гнезд являются кормовые почки, сооружаемые рыжими лесными муравьями, муравьями-древоточцами и некоторыми муравьями-жнецами на дорогах в тоннелях жилого муравейника. Кормовые почки невелики по размерам, в них не бывает молоди, функционируют такие гнезда только летом. В этот период в кормовой почке обитают от нескольких сот до нескольких тысяч фуражиров и строителей.
Временные вспомогательные гнезда облегчают муравьям только использование кормового участка, они не влияют на функции основного муравейника. Невелико значение этих гнезд и для рассредоточения жителей перенаселенного муравейника. Но их роль в удержании территории и более рациональном ее использовании существенна.
От старой сосны, в сгнившей сердцевине которой поселилась семья муравьев-древоточцев, в разные стороны расходятся тоннели. Чтобы обнаружить их, достаточно приподнять слой подстилки из опавшей хвои, тоннели могут быть довольно длинными, до 10-15 м. На разном удалении от гнезда из тоннелей имеются выходы на поверхность. Отсюда отправляются на охоту фуражиры. Заканчиваются тоннели в кормовых почках. Каждое гнездышко так же, как и сам муравейник, размещается в отмершей древесине: пнях, лежащих стволах деревьев. Они и служат опорными пунктами. Из кормовых почек действует основная масса фуражиров. Сюда же поступает большая часть добычи. При большом количестве добычи она поначалу накапливается в почках, а уж затем постепенно переносится в муравейник.
У многих муравьев отработана четкая эстафетная транспортировка пищи: одни рабочие приносят добычу в кормовую почку или к выходу из тоннеля, другие - транспортируют ее оттуда в муравейники. Эстафета экономична: она позволяет обходиться на участке минимумом опытных фуражиров. А транспортировку добычи по тоннелю может осуществить любой.
Большинство кормовых почек остаются неизменно в этом качестве. Но некоторые из них в дальнейшем служат основой для сооружения настоящего муравейника. В специальной литературе описано преобразование в жилые гнезда кормовых почек у рыжих лесных муравьев и пахучего муравья-древоточца. У формика это обычно связано с основанием нового отводка, а у пахучего муравья-древоточца - с разделением семьи.
Другой вариант поликалии связан с разделением гнезд на летние и зимние. Зимние - обычно более крупные, капитальные, с мощной подземной системой ходов и камер. Они расположены в укрытых местах: в пнях, в середине кустов и т. п. Летние гнезда находятся на открытой местности. Они служат муравьям в течение всего периода активности, а в зимние семья перебирается поздней осенью (на зиму в летних гнездах остаются единичные особи).
Летние и зимние гнезда были детально описаны на примере кроваво-красного муравья-рабовладельца одним из пионеров изучения поведения муравьев - Э.Васманом в 1910 г. Сходная картина прослежена у некоторых других видов. Летних гнезд, жилых и вспомогательных, обычно больше, чем зимних. Как правило, концентрация особей в зимовочных гнездах всегда выше. Но из всякого правила бывают исключения. Изучая в Финляндии биологию черноголового муравья, Р. Розенгрен установил, что у этого вида все происходит наоборот: летом муравьи обитают в большом муравейнике, а зимуют вне основного гнезда в небольших зимовочных камерах, сооружаемых вокруг муравейника на различном удалении от него.
Более сложные варианты поликалии возникают при единовременном использовании семьей нескольких жилых гнезд, одного - центрального и нескольких - периферийных. В этом случае отдельные гнезда несут различные функции. Сам факт разделения функций гнезд в поликалическом муравейнике не удивителен, ведь подобное разделение складывается уже при монокалии, т. е. при обитании семьи в одном гнезде. Но там различные функции имеют отдельные части одного гнезда (например, внутренний конус и гнездовой вал у рыжих лесных муравьев). Здесь же обособление частей гнезда пошло дальше и вылилось в принцип: гнездо - функция. Сравним единое сложное гнездо пятнистого муравья с поликалической системой муравьев-портных (рис. 11).
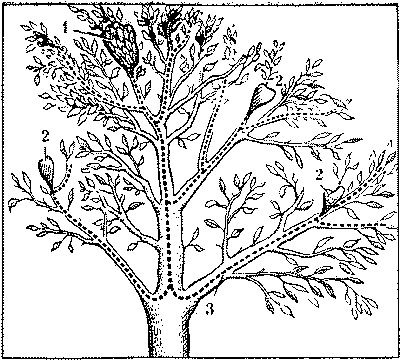
Рис. 11. Цоликалическии муравейник муравьев-портных на дереве
1 - центральное гнездо с самкой, 2 - периферийные гнезда, 3 - дороги
У пятнистого муравья гнездо состоит из трех частей: центрального гнезда, где живет единственная самка и ее свита; большого внутреннего гнезда (сферы), где находятся только рабочие - имаго; периферийного гнезда {сферы) с рабочими и молодью. В центральном гнезде муравьев-портных также находится самка и ее свита. Личинок младших возрастов уносят в разбросанные по кроне дерева периферийные гнезда, где их выращивают до взрослого состояния. Есть также небольшие гнезда, в которых находятся одни рабочие.
В обоих случаях добыча доставляется фуражирами сначала в периферийные гнезда (или сферу) и лишь оттуда поступает в центральное гнездо. Этот принцип сохраняется у всех муравьев, перешедших к поликалической форме существования. Естественно, что между гнездами имеются оживленные дороги и постоянный обмен. Из центрального гнезда в периферийные идет пополнение молодью, обратно - поступают взрослые муравьи, а также пища. Функциональная цельность такого муравейника не вызывает сомнений.
Поликалия позволила муравьям увеличить численность общины и размер используемого кормового участка. Вся территория внутри кольца вспомогательных и периферийных гнезд используется одной семьей даже в тех случаях, когда в других частях кормового участка статус охраняемой территории отсутствует. В этом одно из преимуществ поликалии.
Вообще с переходом к поликалии конкурентоспособность семьи значительно возрастает. Одновременно муравьям гораздо легче приспособиться к среде: гнезда располагаются в разных условиях, появляется возможность маневра при изменении погоды или режима освещенности отдельных гнезд. Виды, освоившие поликалическую форму существования, смогли подняться на довольно высокий уровень численности населения семьи. Подсчет особей в семье муравьев-портных, произведенный Ф. Вандер-планком в 1960 г., показал, что в крупном поликалическом муравейнике обитает более 100 тыс. особей. По данным московского энтомолога А.М. Кравченко, численность семьи пахучего муравья-древоточца еще выше - более 300 тыс. особей.
Уровень численности особей в семье, которого достигают муравьи, освоившие очередной этап развития социальной организации, убедительнее всего показывает адаптивную ценность того или иного достижения. Ряд поликалических видов преодолел стотысячный рубеж численности, т. е. достиг и даже превзошел уровень, при котором у муравьев с наиболее многочисленными семьями (рыжие лесные муравьи) происходит образование колонн.
Это доказывает, что, хотя поликалия располагает несколько меньшими возможностями, нежели система колонн, она оказалась очень важным шагом в эволюции социального образа жизни у муравьев.
Диффузные гнезда можно рассматривать как один из вариантов поликалии, связанный с использованием муравьями готовых полостей и камер, небольшие размеры которых ограничивают возможное число обитающих в них особей. Как правило, диффузные гнезда встречаются у муравьев, особи которых имеют небольшие размеры. Семья как бы растекается по всевозможным укрытиям, легко меняет их, сохраняя при этом связи между отдельными частями. Вместе с рабочими по микрогнездам рассредоточены и самки. Семьи таких видов имеют необыкновенно пластичную, подвижную структуру. Они легко распадаются, и обитатели каждого микрогнезда могут стать основой новой большой семьи. Состав и численность населения микрогнезд могут меняться: увеличивается размер гнезда - там начинает собираться больше муравьев, при необходимости сменить жилище группировка может разделиться на несколько, чтобы потом вновь собраться в следующей обители.
Численность семьи бывает значительной - до 10 тыс. и более. Среди муравьев, обитающих таким образом, наиболее известен фараонов муравей, доставляющий массу хлопот санитарно-эпидемиологической службе у нас и за рубежом. Фараонов муравей ведет настоящее наступление на жилище человека, проникает в библиотеки и на предприятия пищевой промышленности. В больницах он способен совершить настоящие преступления, нарушая стерильность операционных и разнося инфекцию. Борьба с этим крошечным (около 2 мм) желтоватым муравьишкой крайне затруднена именно из-за его особенностей, обусловленных диффузно-кочевым образом жизни. Едва заметные щели, забытая корка хлеба, корешок книги - все пригодно для поселения фараонова муравья. Просачиваясь из квартиры в квартиру, из палаты в палату, он очень быстро осваивает различные помещения, постепенно становясь одним из самых распространенных и нежелательных спутников человека.
Надо сказать, что роль муравьев с диффузными гнездами в природе невелика. Они заселяют брошенные ходы жуков, трещины, плоды и даже шипы некоторых растений и т.п. Впрочем, приспособленность к жизни в чужих жилищах пригодилась не только фараонову муравью и не только в жилище человека. Семьи столь же мелкого по размерам блестящего муравья-малютки отлично приспособились к обитанию в гнездах рыжих лесных муравьев. Крошечные камерки размещаются в простенках между камерами гнезд муравьев-хозяев. Оттуда по ходам диаметром менее 1 мм муравьи-малютки похищают добычу хозяев и даже их яйца и личинок. И всюду, где есть гнезда рыжих лесных муравьев, встречается вид-приживалка. Муравьи этого вида уже не могут жить самостоятельно.
Муравьи с диффузными семьями имеют перед остальными муравьями то преимущество, что им не надо строить жилища. И поэтому их пора наступает тогда, когда появляются чужие жилища, где тепло и всегда есть корм.
Как мы убедились, муравьи использовали разнообразные приспособления, открывающие дополнительные возможности роста семьи за счет усложнения ее организационной структуры. В общих чертах смысл таких приспособлений сводится к решению одной задачи - формированию внутрисемейных организационных единиц и функциональной дифференциации самого гнезда. Соотношение этих составляющих различно в рассмотренных вариантах, различен их эффект. Но все происходит внутри одной семьи. Многие виды ограничились этими достижениями. Следующий этап развития социального образа жизни у муравьев - выход на подсемейный уровень организации. Предпосылки к возникновению подсемейных структур сложились в процессе развития организационной структуры отдельной семьи муравьев. Возможности их образования - в механизмах, регулирующих жизнь муравьиной общины.