3. МУРАВЕЙНИК И КОЛОНИЯ
ОГЛАВЛЕНИЕ:
В предыдущей главе мы подошли к тому моменту в жизни семьи муравьев, когда внутренние возможности общины отрегулировать свой состав и организационную структуру исчерпаны. Целостность семьи под угрозой. В этой критической ситуации происходит деление общины. Тут возможны два варианта: либо семья делится на две равные, сразу же обособляющиеся и становящиеся чужими части (гесмозис), либо из муравейника уходит небольшая часть населения и основывает отводок (почкование). Именно второй ведет к образованию подсемейных структур.
Дело в том, что отпочковавшаяся часть - дочерняя семья - еще долгое время не теряет связи с материнским муравейником. Эта часть организационно оформлена: в ней есть свои самки и рабочие муравьи всех функциональных групп. Появляется у нее и свое гнездо, но связи между старым и новым муравейниками сохраняются. В итоге возникает система родственных муравейников, объединяемых обменными дорогами и определенным типом отношений.
Образование отводка происходит обычно в определенные для каждого вида сроки. Вот как происходил начальный этап выделения нового муравейника у волосистого лесного муравья в 1967 г. в подмосковном ельнике (Солнечногорский лесокомбинат). В течение нескольких дней (15-21 сентября) муравьи проявляли повышенную активность. Огромное количество рабочих возбужденно перемещалось по куполу гнезда и на участке. Особо оживленно было в зоне одной из дорог, на которой предстояло сооружение отводка.
В разных местах кормового участка этой дороги собирались группы рабочих, от нескольких десятков до нескольких сот муравьев; 21 сентября под березой, что росла в 12 м от гнезда, началось сооружение нового муравейника, сначала укрывавшегося под сухими листьями.
Через несколько дней листва была покрыта небольшим плоским куполом из хвои диаметром около 15 см. Вокруг купола появился небольшой вал из свежей почвы:
муравьи сооружали подземные гнездовые помещения. Это были подготовительные работы, а само переселение началось 22 сентября. В тот день с утра на куполе материнского муравейника появилось множество крайне возбужденных рабочих особей, в основном крупных размеров. К 11 часам на поверхность купола вышло несколько десятков самок, каждая из которых была. окружена толпой снующих рабочих.
Измерение муравьев из свиты самок показало, что эти муравьи значительно крупнее рабочих, находящихся на поверхности купола в обычное время. Рабочие непрестанно облизывали медленно передвигавшихся по вершине гнезда самок.
Тем временем муравьи-носильщики начали курсировать между сооружаемым отводком и материнским муравейником и переносить в отводок находящихся на куполе взрослого гнезда самок и рабочих из их окружения. Образовалась широкая дорога, по которой в отводок непрерывным потоком поступали самки и рабочие особи. Рабочие, которых переносили носильщики, были внутригнездовыми с характерным для этой категории особей раздувшимся из-за увеличенного жирового тела брюшком. Ежеминутно в отводок доставлялось 25-30 особей. Фуражиры переходили туда самостоятельно. На следующий день картина повторилась. Затем возбуждение на куполе взрослого муравейника улеглось, но массовое переселение в отводок продолжалось до середины октября.
Переселенцы благополучно зимовали во вновь отстроенном гнезде, достигшем к этому времени 30 см в диаметре и 15 см в высоту.
Отводки основываются на различных расстояниях от материнского гнезда. У рыжих лесных муравьев удаление отводка колеблется в пределах от 10 до 120 м. Дочерние семьи кроваво-красного муравья-рабовладельца поселяются в 15-30 м от материнского гнезда, у тонкоголовых муравьев - в 5-25, у муравьев-жнецов - в 15-20, у муравьев-фаэтончиков в Туркмении - в 10-25 м.
В результате почкования возникает новое образование - колония, состоящая из материнского и одного или нескольких дочерних муравейников. Семьи, входящие в одну колонию, связаны дорогами, по которым производятся обмены. Эти дороги называются в отличие от кормовых дорог муравьев обменными дорогами, что отражает их основное назначение и функцию в колонии. Как уже говорилось, именно благодаря обменам и появилась возможность сохранения в течение длительного времени особых лояльных, так называемых колониальных отношений между муравейниками.
Почкование - не единственный способ возникновения колонии. Довольно часто колония появляется при установлении колониальных отношений между уже существующими семьями разного происхождения, но поселившихся по соседству. Происходит породнение чужих семей муравьев одного вида, что впервые описано Ц.И. Мариковским в 1962 г. Только таким путем формируются искусственные колонии рыжих лесных муравьев. Нами выяснено, что при породнении чужих семей в естественных и искусственных популяциях рыжих лесных муравьев связь между такими семьями устанавливается с помощью небольшого специального гнезда - промежуточной почки, сооружаемой на границе кормовых участков муравейников. В отдельных случаях на дороге, соединяющей соседние гнезда, строятся сразу две промежуточные почки. С обеих сторон к промежуточной почке подходят обменные дороги, по которым во встречных направлениях переносятся самки, рабочие муравьи и молодь. Некоторое время у такой почки существует небольшое население из одних рабочих особей. Значительная часть обменов производится здесь, а муравьи-носильщики, пришедшие на купол чужого гнезда, встречаются там довольно враждебно.
Через две-четыре недели, после проведения большого числа обменов, промежуточную почку покидают, и между породнившимися семьями устанавливается прямая связь. Но если дороги пересекались здесь под каким-то углом, то и после установления прямой связи обменная дорога долго еще сохраняет первичные изгибы, указывающие на места соприкосновения охраняемых территорий различных семей или колоний, связанных вторичными обменными дорогами. Поэтому вместе с другими показателями дорожная сеть может таким образом использоваться при анализе структуры и происхождения объединений муравьиных семей.
При породнении чужих семей бросаются в глаза два непременных условия, соблюдаемые в этом случае муравьями. Во-первых, породниться таким путем могут только чужие в данный момент семьи. Во-вторых, муравейники должны значительно отличаться по своей численности, Суть возникновения колонии посредством породнения чужих семей заключается в вовлечении более сильным, процветающим муравейником ослабевшего соседа в свою обменную систему.
Опять обмены! Обмены пищей между индивидами в колонне (семье). Обмен особями между колоннами одной семьи. Обмены - средство сохранения дружественных отношений между родичами и вовлечение в круг родичей новых особей и целых муравейников.
Ради чего это происходит? Только ради возможности построить побольше гнездо, в котором легче регулировать гигротермический режим? Это объяснение можно принять во внимание, если речь идет о концентрации муравьев в одном гнезде. Но ведь сейчас перед нами другая ситуация: оба муравейника остаются на своих местах.
Неужели колония возникает на тот крайний случай, когда в какой-нибудь черный для муравьев год семьи настолько ослабевают, что только слияние их в одну спасает уцелевших от гибели в пустеющих гнездах? Подобные слияния нередки.
Но причина в другом. Стратегия муравьев заключается в максимальном увеличении числа индивидов в системе, удлинении обменной цепи. Ведь чем она длинней, тем устойчивей система. А пространственная разобщенность семей в колонии при этом не так важна: уже при переходе к поликалии муравьи решили эту проблему даже без нарушения целостности семьи.
И если колониальные связи не устанавливаются между всеми соседними муравейниками, то, по-видимому, лишь потому, что имеется ряд других мощных и препятствующих процессу консолидации факторов. Снять действие этих факторов или хотя бы ослабить их противодействие - и этакий панмуравейник в пределах, например, лесного массива, пожалуй, мог бы стать реальностью. Впрочем, как мы увидим ниже, при определенных условиях такая возможность частично реализуется.
Некоторый из препятствующих объединению факторов уже обсуждались: прежде всего специфический гнездовой запах, четко делящий всех муравьев на своих и чужих. Этот барьер очень высок. Но все-таки он преодолим с помощью интенсивных обменов и небольшого временного гнезда диаметром 15-30 см и высотою 5- 7 см, без подземной части и дифференциации купола. Такое гнездо рыжие лесные муравьи могут построить за день. Очевидно, необходимо еще что-то.
Может быть, влияют размеры муравейников, абсолютные и относительные? Здесь еще многое неясно, но значение этого показателя, несомненно, велико. Ведь недаром для образования колонии требуется, чтобы семьи различались по численности. Попробуем оценить этот фактор в следующих разделах данной главы. А пока внимательно поглядим на первое условие породнения чужих муравейников. Они должны быть чужими, т. е. не входить в одну обменную систему. Но почему обмен, порою весьма слабый, особями налагает вето на установление прямых связей между расположенными рядом гнездами? Не потому ли, что они и так уже входят в одну обменную систему? Если у одного из соседей связь с материнским гнездом оборвется, тогда прямые связи устанавливаются. Такие случаи отмечены в естественных комплексах муравейников. Чтобы яснее представить значение обменов особями для муравьев, возможно подробнее разберем их роль в жизни колонии.
Начальный этап в жизни колонии - просто большое переселение в довольно короткий срок, измеряемый у одних видов днями, у других - неделями. В это время из материнского муравейника поступает в отводок и пища. Дочерняя семья - фактически лишь часть материнской семьи, у которой нет ни собственного охраняемого участка, ни возможности выращивать молодь. Она еще ничего не дает в обмен на получаемых от материнской особей и пищу. Постепенно численность дочерней семьи растет, и характер ее отношений с материнской начинает меняться. На обменной дороге появляются муравьи-носильщики, доставляющие особей из отводка в материнское гнездо. Индивидуальным мочением носильщиков в колониях северного лесного муравья в Подмосковье и муравья-фаэтончика в Туркмении было установлено, что эти особи выходят налегке из своего гнезда и направляются а другое, где забирают объекты обмена (личинок, куколок или внутригнездовых рабочих) и переносят их в свой муравейник. Учитывая схему действия носильщиков, можно считать, что начало обоюдных обменов означает перелом в отношении материнской семьи к дочерней. Это как бы признание отводка отдельным муравейником. Признание еще далеко не полное.
Отводок обычно сооружается очень энергично. У фаэтончиков и муравьев-жнецов уже через две-три недели он имеет все основные элементы гнезда. У рыжих лесных муравьев через месяц-полтора отводок также принимает вид вполне законченного гнезда с внутренним конусом и подземной частью. Это позволяет поддерживать необходимый для развития молоди температурный минимум. Дальнейшие отношения между материнской и дочерней семьями будут зависеть от соотношения их сил (численности) и продуктивности. Сначала наступает период интенсивных взаимных обменов. У муравьев формикаон совпадает с достижением семьей размеров автономного муравейника.
В это время у растущего муравейника начинает складываться и свой кормовой участок. Он еще маленький, проницаем для фуражиров материнской семьи, которые «по старой памяти» продолжают здесь охотиться. Ведь в абсолютном большинстве случаев отводок сооружается в пределах ее охраняемой территории, где-нибудь в периферийной зоне одной из дорог.
Обмен обычно почти равный. На этом этапе в какой-то мере еще выдерживается обменный принцип колонн в гнезде - один к одному. Но появляются и нарастают различия. Материнский муравейник, как более многочисленный, с лучшей терморегуляцией, раньше начинает фуражировку утром, и его носильщики раньше приходят в отводок. А прекращают свою работу носильщики материнской семьи позже носильщиков дочерней. В итога утром и вечером каждого дня материнская семья собирает некоторую «дань» с дочерней. Днем обмен выравнивается, но по итогам суток материнская семья всегда в выигрыше.
Точно так же обстоит дело и в зависимости от сезона. Весной и осенью материнская семья получает гораздо больше (нередко в несколько раз), чем отдает. Положение таким образом изменилось в принципе: отводок служит источником дополнительных особей для материнского муравейника. Но, учитывая небольшие размеры дочерней семьи, в абсолютных величинах прибыль эта невелика.
Весенне-осенние обмены могут быть очень интенсивными, чем, по-видимому, и объясняется рассмотренное выше положение, что именно в эти периоды решается вопрос, быть или не быть отводку. Если дочерняя семья после зимы или неблагоприятного лета оказывается к урочному моменту слишком маленькой, неравный обмен настолько истощает ее, что оставшихся в гнезде муравьев уже недостаточно для жизни отдельной семьей. И тогда ее остатки просто возвращаются в материнскую общину. Таким образом отводок в какой-то мере является резервом материнского муравейника. Процесс разделения общины в определенных условиях обратим. Это еще одно важное отличие почкования от гесмозиса у пчел и некоторых муравьев, о чем говорилось в предыдущей главе.
При благоприятных условиях соотношение сил муравейников в колонии постепенно меняется в пользу дочернего. В период интенсивного выделения отводков материнская семья растет незначительно, поэтому изменения соотношения целиком зависят от роста дочернего муравейника. А он обычно растет довольно быстро, несмотря на невыгодный для него обмен с материнским муравейником. Конечно, при условии, что на кормовом участке достаточно пищи и имеется свободная территория, которую можно освоить.
С ростом отводка интенсивность обменов л колонии начинает падать. Для колонии это имеет очень серьезные последствия. Смысл постоянных обменов между муравейниками заключается в том, что обмены позволяют сглаживать качественные различия между обменивающимися семьями, поддерживать дружественные отношения и избегать столкновений. А сглаживать есть что. Обитание дочерней семьи в новом гнезде и пища, добываемая в определенном участке, приводят к изменению семейного запаха отводка, вначале совпадавшего с запахом материнского гнезда. Чем сильнее отличаются условия обитания и кормовые участки материнской и дочерней семей, тем быстрее накапливаются различия - причина отчуждения муравейников. Пока отводок молод и обмены интенсивны, различия минимальны: С ростом дочерней семьи нивелирующая роль обменов даже при постоянной интенсивности снижается, что ведет к снижению интенсивности обменов, а это, в свою очередь, - к ускоренному накоплению фототипических различий. И чем энергичнее растет отводок, тем это происходит быстрее.
Обмены только пищей и нерегулярные весенне-осенние обмены наблюдаются на последнем этапе существования колонии, предшествующем полному отделению дочернего муравейника. Колонии на этом этапе описаны П.И.Мариковским и мирмекологом из ФРГ Г. Кнайтпем.
Наиболее устойчивым оказывается обмен пищей. Впрочем, строго говоря, поступление добычи в материнское гнездо из отводка нельзя назвать обменом по двум причинам. Во-первых, это процесс односторонний, только в материнское гнездо. Как только отводок подрастает настолько, что начинаются взаимные переносы особей, подкармливайте его со стороны материнской семьи прекращается. Далее пищевой поток направлен только к материнскому гнезду. Получается еще одна форма «дани».
Правда, дань эта собирается не столько в самом отводке, сколько на его кормовом участке. Здесь отношения фуражиров из двух гнезд явно неравноправны. Фуражиры из материнской семьи почти до конца существования колонии могут проникать на кормовой участок дочерней, хотя он уже давно надежно охраняется от всех других соседей. Постепенно случаев такого проникновения становится все меньше, но все-таки они имеются. А вот фуражирам отводка путь на кормовой участок материнского муравейника всегда закрыт. Отсюда и возникает вторая неточность термина. Фактически это не обмен и не «дань» с муравейника, а скорее «концессия», по которой фуражиры одного муравейника охотятся на территории другого.
В тех случаях, когда соотношение сил материнской и дочерней семей изменяется очень быстро, обмен особями между ними иссякает в течение одного сезона. На следующий год еще может быть небольшой весенний обмен и поступление из отводка пищи, но часто не происходит и этого. Все заканчивается в один сезон. События развиваются тем быстрее, чем меньше материнский муравейник, т. е. дочерней семье легче с ним сравняться. Колонии становятся сезонными. Такие колонии имеются, например, у муравьев-фаэтончиков, численность семьи которых ограничивается несколькими тысячами индивидов.
Можно считать, что в течение всего срока существования колонии обмены между материнским и дочерним муравейниками подчинены закономерным изменениям. И если принять весь период, от возникновения отводка до его отделения, за единицу, графики обменов совпадут для колоний с различными сроками существования.
Вообще при оценке подобных явлений абсолютное время характеризует процесс, но не определяет его. Даже сезонность не в состоянии регламентировать срок существования колонии. Она лишь предлагает наиболее подходящие моменты для решающего проявления назревающих в колонии процессов.
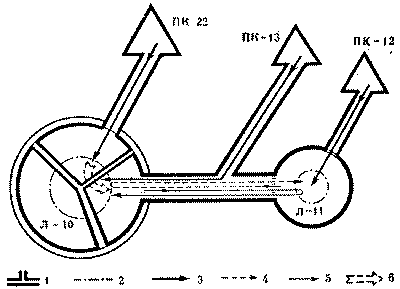
На обмен поступает определенный процент новых рабочих-имаго и молоди, появившихся в гнезде. Но каков процент, сколь он велик? Чтобы проверить, какая часть обменного материала, имеющегося в материнском гнезде и в отводке, проходит на обмен, мы поставили опыты в колонии волосистого лесного муравья. Схема обменов в колонии дана на рис. 12. Крупный материнский муравейник (Л-10) с куполом 2,2 м в диаметре и высотой гнезда 1,1 м имел один отводок (Л-11) с диаметром купола 1,2 м и несколько кормовых почек на разных дорогах. Всего у муравейника Л-10 было шесть дорог, из которых на рисунке изображены только две, использованные в опытах.
Опыты проводились в августе, когда обе подопытные семьи уже вывели последнее поколение рабочих и обменивались только имаго, во сам обмен производился еще в соотношении 1:1. Как видно из схемы, дочерний муравейник и его кормовая почка ПК-12 находятся на одной дороге материнского гнезда. Кормовая почка материнского муравейника (ПК-13) расположена на ответвлении той же дороги. Вторая кормовая почка материнского муравейника (ПК-22) находится на самостоятельной дороге, т. е. принадлежит другой колонне.
В качестве обменного материала использовались куколки, взятые из постороннего муравейника того же вида. Куколки были сосчитаны и окрашены разными цветами. В первом опыте по 2000 куколок выложили на ПК-12 и ПК-13, т. е. на ответвления одной дороги материнского муравейника. После этого подсчитывали поступление меченых куколок из кормовых почек в материнское гнездо и отводок, а на обменной дороге регистрировали взаимную передачу этих куколок жилыми муравейниками.
Обмен начался через 40-45 минут после поступления первых куколок в муравейники и продолжался до конца дня. При этом отводок передал в материнский муравейник 5,6% куколок, а материнская семья - лишь 1,8%, или а 3,1 раза меньше. Причем все время общий обмен между семьями (с учетом продолжавшихся весь день взаимных переносов внутригнездовых рабочих) оставался равным. Этот опыт, повторенный по той же схеме, дал одинаковые результаты. Фактически мы анализировали обмен в пределах колонны, так как и отводок, и обе использованные в опытах ПК находятся на ответвлениях одной дороги материнского гнезда.
Интересно знать, как и в какой мере участвуют в подобных обменах другие колонны материнского муравейника. Для этого в следующих опытах куколок выкладывали на ПК-13 и ПК-22. На обменной дороге этих куколок учитывали по отдельности.
Специфика опыта заключалась в следующем. Куколки из ПК-13 сразу же попадали в место, откуда носильщики из отводка забирали объекты обмена, а куколки из ПК-22 должны были сначала попасть в эту зону из другого сектора, другой колонны. Последнее обстоятельство сразу же сказалось на результатах. Носильщики из отводка, как и в предыдущих случаях, забрали причитающееся им количество куколок. Но доли куколок, поступивших из ПК-13 и ПК-22, отличались в 5 раз: 1,6% - из ПК-13 и 0,32% - из ПК-22. В следующем опыте отличие было шестикратным, т. е. даже в ходе довольно острого эксперимента лишь 15-20% новых куколок из соседней колонны оказалось вовлеченным в обмен с отводком. Из общего числа куколок менее трети процента пошло на обмен. Все это показывает, что связи между отводком и материнским муравейником - это прежде всего связи между отводком и колонной, из состава которой выделилась дочерняя семья. Для остальных колонн материнского муравейника отводок существует как часть этой колонны. Но для отводка наличие и мощность других колонн в материнском муравейнике значимы.
Pис. 12. Фрагмент схемы межсекторного и внутрисекторного обменов куколками в колонии волосистого лесного муравья
Л-10 -
материнское гнездо, Л-11 - отводок, ПК-22, ПК-13,
ПК-12 - кормовые почки;
1 - границы секторов материнского гнезда,
2 - границы внутреннего конуса, 3 -
пути поступления куколок из кормовых почек,
4 - маршруты муравьев-носильщиков из
отводка, 5 - маршруты муравьев-носильщиков
из материнского гнезда, 6 - передача куколок
из сектора в сектор
Так или иначе процент обмениваемых муравейниками новых особей невелик, хотя существен для отводка. Чтобы выдержать равный обмен, он должен жертвовать относительно большей частью своего населения. Данная ситуация важна для отводка, особенно если он еще мал по размерам. Ведь можно предполагать, что обменные возможности каждой семьи не беспредельны. Община, разменявшая налево и направо слишком большую часть своего населения, просто утратит свою индивидуальность, целостность. Какой-то предел должен быть. То максимальное число особей, которое может единовременно обменять семья, сохраняя индивидуальность, будет обменным фондом семьи. Получив новое качество - в виде прошедших линьку личинок, куколок или вышедших из куколок рабочих, семья передает партнеру по обмену определенный процент, соответствующий степени родства между данными муравейниками и отношению их размеров.
Обменный фонд, видимо, составляет небольшую часть населения муравейника. В муравейник северного лесного муравья с населением около 15 тыс, особей была внесена тысяча помеченных групповой меткой рабочих из отдаленной колонии. Пришельцы были приняты враждебно, часть из них была убита, но они все-таки остались в гнезде, влившись в различные функциональные группы местных рабочих. Через день в то же гнездо была выпущена еще тысяча рабочих из третьего, не связанного с первыми двумя, гнезда. Эти рабочие имели новую групповую метку. Их приняли возбужденно, но схваток со смертельным исходом не было. Новые рабочие также влились в состав семьи. И, наконец, еще через день на купол подопытного гнезда выпустили очередную группу рабочих из четвертого, чужого всем предыдущим, гнезда, помеченных своей групповой меткой. На этот раз новички были приняты почти равнодушно. Предыдущие массовые введения новых качеств настолько расшатали монолитность небольшой муравьиной семьи, что она оказалась не в состоянии отличить чужих от своих. Число введенных особей оказалось большим, чем обменные возможности семьи, и она на какое-то время утратила индивидуальность.
Для контроля рабочих из четвертого гнезда выпускали на муравейник из одной колонии с подопытным. Здесь их приняли враждебно. Временная утрата индивидуальности происходит и при формировании искусственной колонии.
Формирование группировок отводков связано здесь с такими массовыми переходами и обменами, что некоторое время после этого уходит на стабилизацию семей. Нестабилизированные семьи легко, без посредства промежуточных почек, объединяются в колонии. В утрате индивидуальности, видимо, еще одна из основных причин поглощения слишком слабых дочерних семей материнскими.
Мы рассмотрели ряд сторон взаимодействия муравейников, входящих в одну колонию. И во всех случаях со всей определенностью проявляется неравноправность этих отношений. Материнская семья выступает как доминант. Доминирование материнской семьи вытекает из самого способа возникновения колонии - почкования. Дочерняя семья, поначалу несравнимо меньшая, может противопоставить этому только свой относительно быстрый рост.
Казалось бы, что с усилением отводка положение может измениться: он вырастет и в колонии установятся отношения, сходные с отношениями колонн в муравейнике. А поскольку отводок может и перегнать материнский муравейник по численности населения, не исключена возможность и смены доминанта. Но этого, как мы видели, не происходит. Обмены между семьями начинают угасать, и накануне выравнивания сил двух муравейников колония распадается. Семьи становятся друг другу чужими, между ними теперь могут начаться даже войны. Ни до координационной системы, ни тем более до смены доминантов дело не доходит. Впрочем, в некоторых случаях, когда в результате разрушения материнский муравейник резко слабеет, часть его населения может перебраться в сохранившийся отводок. Тогда оставшаяся в материнском гнезде община может попасть в зависимое положение. Но такая ситуация и крайне редкая и неестественная. Сам же принцип возникновения колонии и характер отношений входящих в ее состав муравейников показывают: колония - система иерархическая, в которой обязательно доминирует материнская семья.
Основа ее доминирования - большая масса этой семьи. Именно соотношение масс и определяет в рамках вида исход любого события в жизни муравейников.
Но неужели разрыв выровнявшихся по силе муравейников неизбежен? Ведь в его основе, насколько сегодня можно судить, лежит все-таки не столько само равенство, сколько стремительно нарастающее в процессе выравнивания семей расхождение их по фенотипу. Муравьям становится все труднее признавать друг друга своими. И отчуждение происходит из-за того, что в каждом гнезде, прежде всего в отводке, слишком быстро накапливается новое качество. Столь быстро, что обмены не в состоянии сделать его общим достоянием, выровнять семьи по этому признаку до нужного уровня.
А если накопление различий идет так медленно, что обмены в состоянии полностью компенсировать их? Тогда колония сможет существовать неопределенно долго. Теоретически такой вариант возможен. А практически? Длительные наблюдения за многочисленными гнездами рыжих лесных муравьев показывают, что иногда данная тенденция проявляется, причем скорее в тех случаях, когда кормовые и территориальные возможности биотопов не допускают быстрого роста муравейников. Процесс отделения отводка может затянуться на 10 и более лет. Однако в конечном счете разрыв связей происходит, и дочерний муравейник становится самостоятельным. Выход отводка из колонии, по-видимому, правильно трактовать именно так, поскольку обмены объективно направлены на удержание отводка в составе колонии, противодействуют отделению дочерней семьи. Оно происходит не при достижении какого-то определенного, специфичного для вида уровня численности, а после приближения размеров отводка к размерам материнского муравейника.
Чем больше материнский муравейник, тем крупнее должен быть и отводок к моменту отделения. Это обстоятельство очень важно для оценки почкования как способа образования новых семей у муравьев. Естественный отводок в состоянии выделить только взрослый, развитый муравейник. И выйти из-под контроля такого муравейника может муравейник, ставший взрослым и развитым. Чем сильнее материнская семья, тем сильнее, а значит жизнеспособнее будет и отделившийся отводок. Если только позволяет территориальная обстановка, многие отделившиеся отводки в последующие один-два года уже становятся материнскими и выделяют отводки.
Пока что мы говорили о колониях, состоящих всего из двух жилых муравейников, материнского и одного отводка. У видов с малочисленными семьями обычно так и бывает: на формирование нескольких отводков одновременно у материнской семьи нет сил. Но для семей-миллионеров, таких, как у рыжих лесных муравьев, реально образование и удержание с помощью обменов сразу нескольких отводков.
Если у материнского муравейника несколько отводков, они должны быть размещены по какому-то принципу. Этот принцип, как и само образование каждого отводка, должен вытекать из структуры материнской семьи, т. е. любое новое почкование - все тот же способ выравнивания ее колонн, но повторенный несколько раз. Очевидно, что в ряде случаев выделения одной дочерней семьи недостаточно для выравнивания колонн. Тогда отводки должны располагаться на одной дороге материнского муравейника.
Именно так обычно и обстоит дело. Вдоль наиболее сильной дороги материнского гнезда располагаются серии отводков. Помимо отводков в серию могут входить и кормовые почки. Однако у муравейника несоответствие в размерах колонн может складываться не только за счет одной из них. Например, из-за ограничения кормового участка с двух сторон у пятиколонного муравейника две колонны будут постоянны и минимальны по размерам. А три другие, ничем не ограниченные, имеют возможность быстро развиваться. В этих условиях выравнивание происходит за счет всех трех развивающихся колонн. Каждая из них в той или иной последовательности образует дочерние семьи. Тогда вдоль дорог муравейника возникает несколько серий отводков и вспомогательных муравейников. Функциональная схема такой колонии, расположенной в сосняке - зеленомошнике (Приокско-террасный заповедник), дается на рис. 13.
У муравейника Р-49 семь отводков и пять кормовых почек, распределенных по трем дорогам. Отводки разновозрастные и отличаются как но размерам, так и по характеру связи с материнским гнездом. Расположение отводков показывает последовательность их возникновения в пределах одной серии.
Сначала образовались более удаленные отводки 0-3 и 0-6. Они уже настолько крупны, что обменов особями между ними и материнской семьей нет, хотя связующие дороги еще сохраняются. Позже были сооружены гнезда 0-2, 0-4 и 0-7, Они мельче первых и еще обмениваются с Р-49 особями, хотя интенсивность обменов невелика - до 20 переносов в 10 минут.
Отводок 0-5 едва достиг размеров минимального автономного гнезда. Сюда ведет широкая дорога с интенсивными обменами. Наконец, самый маленький - 0-1 находится еще на этапе формирования.
Специфика серийной структуры колонии заключается в том, что каждый отводок, независимо от расстояния и количества гнезд в серии непосредственно связан с материнским муравейником. Это видно и из приведенной схемы колонии. Обмен между отводками одной серии в принципе возможен, особенно при ослаблении их связей с материнским муравейником.
Однажды в колонии волосистого лесного муравья удалось даже наблюдать редкий вариант: отводок, расположенный ближе к материнскому муравейнику, в силу различных причин обогнал в росте более старый и удаленный отводок, расположенный в той же серии. В течение двух лет он отделился от исходной колонии, «прихватив» с собой и отставшего в росте родственника. Отстающий оказался в роли отводка стремительно развившегося молодого муравейника.
А вот муравейники, расположенные на разных дорогах, не могут обмениваться непосредственно друг с другом. Это невозможно даже для гнезд, расположенных на различных ответвлениях одной дороги, например, для отводков 0-1 и 0-2. Такое положение вытекает из схемы работы муравьев-носильщиков, приведенной на рис.12. Обмен между сериями отводков производится только через метрополию, опосредованно. Мы застали колонию Р-49 в момент, предшествующий отделению первых отводков, но пока все еще «в сборе». Естественно, чтобы доминировать в такой многосемейной колонии, материнский муравейник должен быть очень сильным и иметь огромный обменный фонд. Ориентировочная численность семьи Р-49 - около 2,5 млн. особей. В день эта семья передавала на обмен в отводки около 9 тыс. особей, включая личинок, куколок и внутригнездовых рабочих. Данная цифра, по-видимому, максимальна, поскольку сюда входят и переносы во вновь создаваемый отводок 0-1. Но, с другой стороны, ясно, что в близком к зарегистрированному нами режиму муравейник находится уже несколько лет. В отводках 0-3 и 0-6 уже сформировались по три колонны; эти муравейники (1,1-1,2 м в диаметре купола) таковы, что могут сами образовывать отводки. Подобные случаи иногда наблюдаются в природе.
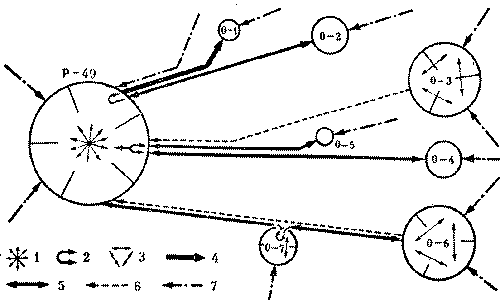
Рис. 13. Схема обменов в колонии обыкновенного муравья (фрагмент) Р-49 - материнский муравейник; 0-3, 0-6, 0-7 - дочерние муравейники с тремя и двумя колоннами; 0-1, 0-2, 0-4, 0-5 - дочерние муравейники с одной колонной; 1 - многосторонние обмены между колоннами материнского муравейника, 2 - эстафетный обмен особями между муравейниками одной серии, 3 - обмены особями между колоннами отводков, 4 - односторонний перенос особей из материнского гнезда в отводок, 5 - двухсторонний обмен особями, 6 - перенос пищи из отводка в материнский муравейник, 7 - поступление пищи с кормовых дорог
Наконец, логично предположить, что крупный материнский муравейник имеет меньшую пропускную способность при обменах между разными сериями в колонии, а следовательно, чем крупнее материнское гнездо, тем меньше родство дочерних семей, относящихся к разным сериям.
Степень родства не зависит от расстояния между гнездами. Размещение отводков и кормовых почек определяется рядом причин. Прежде всего - это расположение и плотность первичных, уже существующих гнезд и кормовых почек. Первые отводки, если нет иных ограничений, распределяются довольно произвольно, часто на самой границе или даже вне охраняемой территории материнского гнезда. Процесс почкования, таким образом, несет еще и функцию расселения, освоения муравьями новых земель. На размещении отводков сказывается наличие конкурирующих муравейников того же или других видов муравьев. В условиях острой территориальной конкуренции серии отводков, кормовые и обменные дороги располагаются параллельно друг другу, что показано на схеме колонии Р-49.
Если территориальной конкуренции нет, дочерняя семья может уйти так далеко от материнского гнезда, что кормовые участки двух муравейников оказываются пространственно разобщенными, т.е. здесь наблюдается прерывистость территории. В изображенной на рис. 14 колонии кормовые участки двух семей северного лесного муравья с самого начала отдалены друг от друга и связаны лишь полосой обменной дороги, которая не имеет других функций. Дочерний муравейник, созданный на небольшом плацдарме вне кормового участка материнского гнезда, в дальнейшем сам осваивает новую территорию. Следовательно, в этом случае образование отводка связано с расселением вида и не ущемляет в дальнейшем интересы материнской семьи. В то же время прерывистость территорий способствует, вероятно, более быстрому отчуждению дочерней семьи.
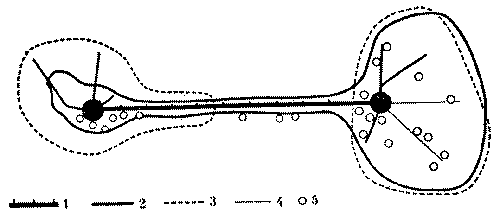
Рис. 14. Развитие колонии северного лесного муравья при отсутствии территориальной конкуренции
1 - обменно-кормовая дорога в 1966 г., 2 - границы охраняемых территорий в августе 1966 г., 3 - границы охраняемых территорий в августе 1967 г., 4 - кормовые дороги, 5 - деревья с колониями тлей
В тех случаях, когда формирование отводков происходит в условиях территориальных ограничений, материнская семья вынуждена создавать отводки на окраинах своего кормового участка. Пока дочерний муравейник мал и не имеет собственной охраняемой территории, его существование не сказывается на экономике материнской семьи. Однако в дальнейшем он начинает вытеснять материнский муравейник из кормовых угодий, прилегающих к отводку. В результате кормовая база ведущей ранее колонны резко сокращается, а наиболее сильной оказывается следующая по численности. Выделенные ею отводки тоже со временем отделяются, «прихватив» куски территории материнского гнезда. После этого наступает черед следующей колонии.
В классическом варианте подобная участь по очереди настигает все колонны, каждая из них теряет часть своего кормового участка. В итоге кормовой участок семьи выравнивается по наименьшей дороге. Процесс выравнивания продолжается до тех пор, пока в распоряжении материнского муравейника не остается совсем маленькая территория, пищевые ресурсы которой не позволяют формирования отводков вообще.
Примерно такую ситуацию мы имеем в колонии волосистого лесного муравья (рис. 15). За 10 лет (1966-1976) в результате поочередного изъятия все новых кусков исходной территории вырастающими отводками материнский муравейник оказался зажатым буквально на пятачке радиусом 15-18 м. Все возможности дальнейшего роста материнской семьи исчерпаны. Теперь, если ситуация радикально не изменится, например в результате случайной гибели одного из отводков, материнская семья обречена. Ограниченные ресурсы оставшегося в ее распоряжении участка недостаточны даже для поддержания населения на имеющемся уровне численности, а, неизбежное уменьшение населения повлечет за собой дальнейшее сокращение участка. Семья оказывается в тисках, которые постепенно сжимаются.
Что же происходит? Продуцируя отводки, муравейник в конечном счете обрекает себя на вырождение, гибель?
Именно так. При многолетних наблюдениях за одними и теми же комплексами гнезд приходится регистрировать этот неумолимый процесс. Процветавшие десятилетие назад муравейники на глазах утрачивают свою былую силу. Вокруг некоторых из них уже сомкнулись границы новых, ими же выделенных когда-то отводков.
Впрочем, положение преемников весьма сложно: они сразу же попали в условия жесткой территориальной конкуренции. Им обычно негде развернуться так, как когда-то это делали обитатели первых гнезд комплекса. Такая ситуация характерна для центральной части комплекса, а по периферии вырастают все новые мощные муравейники, и продолжается освоение еще не захваченных этим видом свободных территорий. До тех пор, пока все пригодные для существования угодья не будут заняты или на пути не появятся владения конкурирующих муравьев, непреодолимые преграды и т. п.
В центральной же части комплекса положение постепенно стабилизируется. Муравейники выравниваются по размерам гнезд и площади кормовых участков. Между соседними семьями устанавливается напряженное динамическое равновесие. В этих условиях всякое ослабление семьи вызывает быстрый натиск соседей, передел территории. С уменьшением одного муравейника увеличивается другой. Переделы территорий нередко сопровождаются войнами, истощающими конкурентов. Радикально решая насущные, сиюминутные задачи по сохранению семьи, муравьи не в состоянии предвидеть и оценить отдаленные последствия почкования для собственной семьи. Да собственно это чисто риторическая постановка вопроса, ибо представить ситуацию в целом некому: ни один из муравьев на это не способен, а специальный координирующий действия колонн центр в семье отсутствует. Согласование, достигаемое почти механическим взаимодействием многотысячных масс муравьев, не позволяет подняться до уровня прогнозирования событий. Перспектива для материнского муравейника оказывается неутешительной. Для отдельного муравейника, но не для популяции, которой обеспечены не только быстрое и эффективное распространение по территории, но и стабильный уровень численности при максимальном использовании ресурсов занимаемой территории.
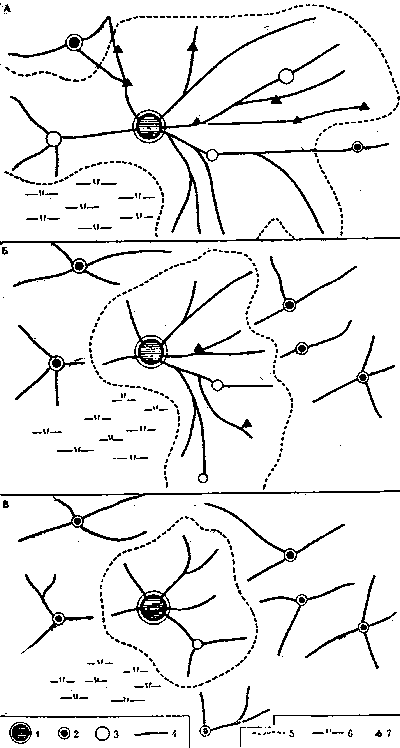
Рис. 15. Сокращение кормового участка материнского муравейника в результате образования и отделения отводков
Положение в колонии в различные годы: А - 1966 г.; Б -1970 г.; В - 1976 г. 1 -материнский муравейник, 2 - отводки, отделившиеся или имеющие охраняемую территорию, 3 - отводки без охраняемой территории, 4 - дороги, 5 - границы кормового участка материнского муравейника, 6 - участок, непригодный для обитания муравьев, 7 - кормовые почки
Все это ожидает материнскую семью впереди. А пока она доминирует в колонии, представляющей из себя семью муравейников: мать и дети. Материнская семья, продуцируя дочерние, сохраняет свою целостность, свою индивидуальность. Каждая дочерняя со временем приобретает свою индивидуальность, в чем-то неповторимую так же, как неповторимо гнездо, в котором она обитает.
Если мы сравним гнезда рыжих лесных муравьев разного возраста, то заметим интересную закономерность. Гнезда-первогодки одного вида удивительно похожи друг на друга. Примерно так же, как похожи суточные цыплята. Форма этих муравейников фактически еще не отражает ни особенностей организации каждого из них, ни условий обитания, в которые попала юная семья. В архитектонике такого гнезда в зачаточной форме заложены все необходимые для последующего развития муравейника элементы, специфичные для этого вида. Конечно, некоторые факторы среды сказываются еще в момент закладки муравейника. Скажем, если гнездо строится на большом пне, то габариты последнего неизбежно отразятся на объеме и габитусе муравейника. Но это особый случай, не отвергающий самого принципа начального стереотипа гнездостроения, проявляющегося на первых порах существования муравейника. В дальнейшем в зависимости от особенностей организации и состояния семьи и условий ее обитания изменяется форма гнезда и соотношение его отдельных элементов. В результате, обследовав тысячи взрослых муравейников, мы не сможем найти двух одинаковых.
Еще оригинальнее по составу, организации и другим показателям обитающие в этих гнездах семьи. Конечно, изучение индивидуальных особенностей муравейников дело многотрудное, да и острой необходимости в этом нет. Но даже сравнив некоторые, лежащие на поверхности характеристики, например структуру кормовых участков различных семей, мы придем к выводу о значительных отличиях в составе каждой из них.
Вопрос индивидуальности и целостности - один из центральных вопросов биологии. Говоря о муравьях с их сложной и развитой «социальностью», мы сталкиваемся не только с выраженной индивидуальностью каждой отдельной особи, но и целых сообществ. Причем индивидуальность семьи или входящей в состав семьи колонны не имеет ничего общего с индивидуальностью организма, ибо базируется целиком на поведенческом взаимодействии составляющих общину индивидов. Сообщество муравьев или других общественных насекомых не теряет специфики как сообщества даже тогда, когда в результате развития внутрисемейных структур оно выходит на уровень, допускающий его деление.
В принципе образование колонии представляет собой растянутый во времени процесс деления муравейника, сохраняющего при этом целостность и индивидуальность. Все это очень наглядно проявляется в упрощенном варианте, когда колония состоит из двух семей - материнской и одного отводка, как, например, показано на рис. 14. Однако мы знаем, что если отпочкование одной дочерней семьи не приводит систему колонн в устойчивое (равновесное) состояние, образуется вторая, третья и последующие дочерние семьи. Время замедляет свой бег, зато отчетливо слышны внутренние часы колонии: спешащие стрелки отводков и сдерживающий их маятник материнского муравейника. В этом тоже проявляется индивидуальность семей. У каждой своя задача: дочерние семьи стремятся подняться к оптимуму, материнская - удержаться в оптимальном режиме.
Здесь уместно вспомнить о диффузных муравейниках. Почему применительно к ним мы не употребляем термин «колония»? Ведь и там несколько гнезд, хоть и маленьких, связаны обменными дорогами. Бывает, что связь между одним из таких гнезд и остальными обрывается. И тогда, что очень важно, этот обособившийся муравейник не погибнет, а начнет развиваться как самостоятельный: в нем, как и в остальных, есть яйцекладущие самки. Чем не почкование?
Да, действительно, внешнее сходство имеется, но только внешнее. Ведь в микрогнездах диффузного муравейника нет главного - целостности и индивидуальности. Население этих гнездышек случайно и непостоянно. Оно, как живая ртуть, перетекает из убежища в убежище. Единственным проявлением целостности здесь является целостность всей семьи. В этих условиях речь идет не о выделении части семьи - части, обеспеченной полным составом и достаточной для самостоятельного существования численностью, а о потере, случайном отщеплении группы особей. Правда, эта группа живуча и начинает вести себя как целая семья. Но и произвольно взятая часть любой колонны, отделенная от муравейника и перенесенная на место, где нет муравьев, способна реорганизоваться и имеет шанс развиваться во взрослый муравейник. Именно эта возможность муравьев к подобной реорганизации и лежит в основе всех методов переселения полезных видов муравьев искусственными отводками. Были бы в таком отводке самки и достаточное число рабочих.
Иногда поговаривают о способности семьи к регенерации при гибели части особей, проводя при этом аналогию с некоторыми растительными или животными организмами. Однако ни о какой регенерации здесь не может быть и речи. С одинаковым успехом можно обсуждать регенерацию средневекового города после чумы. В действительности семья, потерявшая по какой-либо причине (например, взят искусственный отводок) значительную часть особей, со временем восстанавливает частично или полностью свою численность, как восстанавливает ее популяция после депрессии. Искусственно переселенный отводок начинает развиваться на новом месте как совершенно новое образование. И не стоит думать, что если он взят из пятиколонного муравейника, то вырастет именно в пятяколонный.
Итак, колония - образование временное и наделенное определенными функциями. Основной функцией колонии является выращивание новых муравейников. Серийная структура колонии выросла из организационной структуры материнского муравейника. Реальная структура каждой колонии на любом этапе ее существования продолжает отражать прежде всего состояние и структуру материнского муравейника. Значение колонии как эффективного пути размножения семей и расселения муравьев трудно переоценивать. Именно благодаря колониальности ряд видов получил возможность заселять огромные территории и поддерживать стабильную высокую плотность поселения.
Казалось бы, образование колоний, тем более существующих длительное время, - это последняя вершина организационной структуры муравьиного сообщества. Однако, как уже говорилось в предыдущем разделе, колония, как и все прочие организационные достижения муравьев, небезупречна. Длительный процесс образования отводков ведет в конечном счете к перенаселению, а это очень опасно, так как влечет за собой вырождение.
Возможность перенаселения - косвенный результат одного из существенных недостатков иерархического принципа организации колонии. В самой колонии, так же как и в семье, численность особей регулируется в соответствии с пищевыми ресурсами участка. В семьях рабочие муравьи поедают лишних личинок при недостатке пищи. Избыточное число муравьев выселяется в отводки. Рост отводков сдерживается неравными обменами, а рост материнской семьи - последовательным сокращением ее кормового участка. Пока все семьи в колонии, перенаселения не будет.
Но все-таки тенденции к росту отводков столь сильны, что они постепенно высвобождаются из-под регулирующего влияния колонии. Вот тут-то и возникает опасность перенаселения. Инициатива принадлежит одиночным, вышедшим из колоний муравейникам. Каналов регуляции для них стало меньше, а возможностей использования кормового участка больше. Теперь вся без исключения добыча остается в таком гнезде. И если общая численность гнезд в комплексе достаточно велика, продолжающий свой рост отводок быстро достигает предела, обеспечиваемого ресурсами его кормового участка.
Одиночные муравейники являются стихийными элементами, они начинают теснить соседей, захватывая часть их кормовых участков. Начинаются истребительные войны, основным итогом которых, как правило, бывает лишь гибель множества муравьев. Это тоже регуляция численности, но производится она явно нерациональным путем.
Происходит все потому, что выросший отводок должен отделиться от колонии, чтобы не нарушать незыблемого принципа доминирования материнского муравейника. Он должен выпасть из системы надсемейной регуляции численности. Да и сама система обменов в колонии довольно несовершенна. Именно в тот момент, когда регуляция важнее всего, интенсивность обменов стремительно падает. Масштабы колонии, даже самой крупной, тоже недостаточны. Впрочем большего от колонии требовать трудно, ведь ее регуляторные возможности сводятся к возможностям лишь одного материнского муравейника. Таким образом, сам принцип иерархии семей в колонии ограничивает размеры такой системы муравейников и во времени, и по числу входящих в систему семей. Следовательно, для дальнейшего расширения границ системы регуляции численности принцип не пригоден. Но отказаться от него невозможно, ибо колония уже есть, и с ней у ряда видов связана сама возможность появления новых муравейников.
Тем не менее выход есть. Он заключается в новом этапе развития надсемейных структур у муравьев. На этом этапе возникают объединения колоний. Наиболее ярко проявляются такие объединения у северного лесного муравья, ставшего благодаря этому одним из наиболее многочисленных видов наших лесов. Система взаимосвязанных колоний - федерация возникает именно в условиях перенаселения, нехватки охотничьих угодий.
В федерации каждая колония сохраняет свою структуру с доминированием материнского муравейника. Более того, обмены внутри каждой колонии становятся интенсивнее, при этом интенсивность в меньшей степени, чем в отдельной колонии, зависит от соотношения сил муравейников. Это означает, что родственные связи внутри колоний, входящих в федерацию, более прочные и стабильные, а возможности отделения отводков соответственно меньше. Но самое важное - это принцип объединения колоний. Оно должно быть таким, чтобы не затрагивало внутренней структуры составляющих единиц, не разрушало принципа колониальности. Иначе вместо нескольких колоний возникнет одна, состоящая из многих муравейников. А чем больше гнезд в колонии, тем она эфемернее.
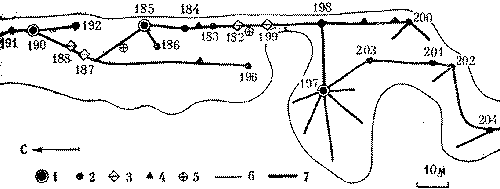
Рис. 16. Федерация муравейников северного лесного муравья (фрагмент)
182-204 - номера гнезд; 1 - гнезда-доминанты, 2 - жилые муравейники, 3 - буферные гнезда, 4 - кормовые почки, 5 - брошенные гнезда, 6 - границы пригодного для использования муравьями участка, 7 - муравьиные дороги
Основная угроза устойчивости федерации заключается в том, что среди материнских семей, вступивших в контакт, выделится сильнейшая, которая и станет доминантой по отношению ко всем, входящим в федерацию семьям. Поэтому прямого противостояния материнских семей надо избегать. Это достигается весьма простым способом: обменная связь материнских муравейников устанавливается только через отводки или специально сооружаемые промежуточные почки (как при породнении чужих гнезд в колонии). Но здесь промежуточные почки действуют постоянно. На рис. 16 показан фрагмент федерации северного лесного муравья, состоящий из 19 жилых муравейников. Видно, как из трех ранее обособленных колоний образовалась федерация. Первая колония включала до объединения четыре муравейника: 190 (материнский), 191, 192, 196 (отводки). Вторая колония состояла из четырех муравейников: 185 (материнский), 183, 184 и 186 (отводки). Третья колония наиболее мощная. Здесь имеется первичный материнский муравейник 197, один вторичный материнский муравейник 198 с отводком 200 и отводки муравейника 197-201, 202, 203 и 204, расположенные на одной дороге этого гнезда. Дорога изгибается, обходя не пригодные для муравьев слишком сырые участки. Принцип соединения колоний внутри федерации показан на рис. 17.
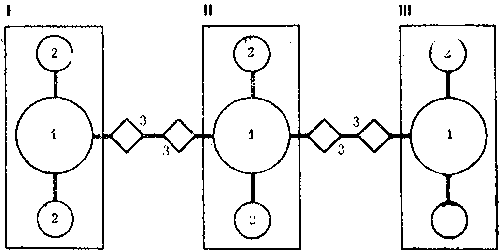
Рис. 17. Блок-схема федерации северного лесного муравья
I, II, III - колонии, входящие в состав федерации. 1 - материнские муравейники, 2 - отводки, 3 - буферные гнезда
Появление вторичных материнских муравейников симптоматично для федерации. Оно показывает, что даже очень крупные отводки, которые в состоянии сами образовывать дочерние муравейники, при интенсивных обменах удерживаются в составе колоний. В нашем примере муравейник 197 имеет диаметр купола 2,2 м, а его отводок 198 - 1,9 м. Обе величины предельны для данного вида. В сравнении с этими гнездами два других материнских муравейника выглядят карликами (диаметр купола гнезда 190 равен 1,2 м, гнезда 185 - 1,3 м). Тем не менее никакой иерархии колоний нет, так как при объединении между ними были сооружены буферные гнезда - постоянные промежуточные почки. Буфером между колониями 1 и 2 служат муравейники 187-188, между колониями 2 и 3 - 182 и 199.
Объединение колоний, по-видимому, не обошлось без предварительных столкновений на территории. Об этом свидетельствуют имеющиеся в пограничных зонах колоний следы старых брошенных небольших гнезд, что обычно указывает на острую территориальную конкуренцию (см. рисунок). Однако теперь сражения за территорию в прошлом, все муравейники оказались связанными общей цепью обменов. Их интенсивность очень высока во всех звеньях федерации. Интенсивность переноски особей на основных дорогах, связывающих гнезда-доминанты, достигают 40 и белее в минуту (до 200). На остальных дорогах интенсивность - 15-30 переносов в минуту, что соответствует максимальным величинам в отдельных колониях. В колонии обмены в 40 и более переносов в минуту отмечены только в период формирования отводка.
Все обмены осуществляются через промежуточные почки.
Называть эти гнезда промежуточными почками можно только по аналогии. Фактически буферные гнезда - это особый тип летних гнезд, выполняющих «федеральные функции». Здесь имеется постоянное, частично смешанное население, среди которого высока доля муравьев-носильщиков. Благодаря этим муравейникам становится возможным эстафетный обмен между материнскими семьями. В обменных операциях участвуют десятки тысяч особей. Часть из них занята поддержанием в порядке буферных гнезд, другие осуществляют переносы.
Каким образом осуществляется запуск этого мощного механизма интеграции, неизвестно. С точки зрения экономики отдельных семей столь интенсивные обмены, по-видимому, мало рациональны. Но тем не менее это происходит, в виде федерации панмуравейник получает свое приближенное воплощение. Если в отдельной семье или колонии социальная регуляция колонн или входящих в колонию муравейников возникает как механизм, обеспечивающий общий рост населения, то в федерации направленность регуляции меняется. Здесь единственная функция обменных операций - ограничение общей численности муравьев в системе. Результатом действий регуляторных механизмов в семье и колонии является увеличение числа подсистем (колонн, семей), расселение вида; результатом регуляции в федерации - сохранение основных структурных единиц. О расселении речь не идет, поскольку федерации возникают в условиях, когда расселяться муравьям уже некуда.
И все это достигается одними и теми же средствами - обменами. Чем жестче территориальная конкуренция, тем интенсивнее обмены.
Данный принцип оправдан не только в федерациях. В эксперименте, проведенном на колонии муравьев-фаэтончиков в Туркмении в 1972 г., искусственное сокращение кормовых участков материнского и дочернего муравейников вызвало увеличение интенсивности обменов в 10-15 раз. Одновременно у подопытных муравейников появилась охраняемая территория, не свойственная фаэтончикам в обычных условиях.
Очень четкое разделение охраняемых территорий всех муравейников наблюдается в федерации. Такая взаимосвязь двух весьма различных по сути явлений понятна, так как оба они вытекают из объективной необходимости более эффективной регуляции в условиях жестких территориальных ограничений.
Сочетание четкого разграничения кормовых участков с интенсивными обменами обеспечивает стабильность федерации как системы муравейников. Устойчивость федерации значительно выше, чем обособленной колонии. Хотя разрыв отдельных связей между колониями возможен, тут же возникают новые, выполняющие ту же функцию. Сами же муравейники почти не меняются. За пять лет наблюдений в рассматриваемой федерации северного лесного муравья не погибло ни одного желтого муравейника и не возникло ни одного нового. Соотношение гнезд по размерам сохранилось приблизительно одинаковое, чего не бывает столь продолжительное время в самостоятельной колонии. Фактически неизменными по интенсивности остались и обмены, производимые муравьями летом.
Наименее устойчивыми элементами в федерации оказываются межколониальные обменные дороги и связанные с ними буферные гнезда. Они могут сдвигаться на местности, но сохраняют при этом полную функциональную преемственность. Таким образом в федерации реализуется стратегическая задача: обеспечение регулирования исключительно поведенческими средствами плотности поселения вида. Регуляция и устойчивость при максимальной для данных условий плотности поселения популяции. Такая задача в мире насекомых оказалась под силу только муравьям, точнее - лишь некоторым их видам. Все муравейники, объединенные в находящиеся под наблюдением федерации, активны, а гнезда - в хорошем состоянии. Почему? Ведь вся система функционирует в напряженном режиме (об этом можно судить хотя бы по интенсивности обменов). Может быть, следует сделать допущение, что именно такой режим и является лучшим средством, предохраняющим муравейники от старения и вырождения. Точнее, единственным шансом сохранить активное состояние в условиях перенаселения. Мы можем проверить это предположение, ознакомившись с развитием муравейников тех видов, у которых есть колонии, но не отмечены федерации. Например, волосистого лесного муравья. Гнезда у этого вида, как правило, крупнее гнезд северного лесного муравья, обычно обилие кормовых почек и отводков, но федерации нет. Собственно и не важно, может ли волосистый муравей создать федерацию в принципе. Достаточно взять какой-либо крупный комплекс, в котором федераций нет, а плотность поселения угрожающе высока. Ведь всякая возможность реализуется не всегда. В частности, старый муравейник, стабилизировавшийся в своем одиночном существовании, не дает отводков и не войдет в федерацию. Даже если это будет муравейник столь склонного к федерациям северного лесного муравья.
Весьма обычен комплекс муравейников, «законсервировавшихся» раньше, чем сложились условия перенаселения, подталкивающие муравьев к объединению в федерации. Но за счет отдельных, разбросанных по комплексу колоний плотность поселения все-таки возрастает и достигает критического уровня. Территориальная конкуренция обостряется, разыгрываются ожесточенные сражения между соседними муравейниками, но за этими сражениями не следует объединение. Периодические сражения - плохой регулятор численности. Вскоре из-за нехватки белковой пищи изменяется пищевой баланс семей (возрастает доля пади), сокращается количество выкармливаемой молоди. Теперь уже не до отводков.
Семьи становятся все нетерпимее к чужим особям, в частности к чужим самкам. Замена старых самок молодыми затрудняется. Семьи приходят в упадок, погибают. От комплекса остаются отдельные, едва влачащие существование муравейники или периферийное кольцо муравейников, из которых со временем начнется повторное заселение центральной части комплекса.
Значит, одной системы охраняемых территорий еще недостаточно, чтобы поддерживать комплекс в активном состоянии долгое время. В комплексе остается стихийное начало, ведущее к подъемам и депрессиям, подобным зигзагам движения численности у одиночных насекомых. Конечно, амплитуды колебаний здесь не столь значительны, да и катастрофические спады случаются очень редко. Но все же время от времени они происходят.
А где-то рядом существуют вполне благополучные федерации.
Следовательно, тенденция к стабилизации, проявляющаяся у муравьев на всех уровнях организации, становится, несомненно, положительной только в тех случаях, когда она связана и со стабильной активностью муравейника. Лишь тогда она в состоянии отодвигать границы допустимой плотности поселения без опасности последующего вырождения.
Надо сказать, что для абсолютного большинства видов насекомых эта задача, по-видимому, вообще невыполнима. Поэтому мы и наблюдаем непрерывные колебания численности различных массовых вредителей и менее массовых видов, не причисленных по этой причине к вредным. Только появление достаточно эффективных способов социальной регуляции допускают стабильность. И чем совершеннее социальная организация вида, тем выше уровень, на котором стабильность сохраняется. Сколь же высок такой уровень для муравьев, добравшихся в ходе эволюции до уровня колоний и федераций?
В комплексах муравейников северного лесного муравья, еще далеких от верхнего предела численности и образования федераций, в среднем на 1 га обитает 5-7 млн. муравьев. Плотность поселения того же вида рассмотренной федерации превышает 15 млн. взрослых особей на 1 га. Весит население такой федерации более 120-130 кг. Это вес стационарный, т. е. постоянно поддерживаемый при имеющемся довольно высоком уровне смертности на участке.
Эти цифры не идут ни в какое сравнение с соответствующими среднегодовыми величинами для любых других насекомых.
Федерация, пожалуй, последнее, высшее достижение социальной организации у муравьев. Прослеживая восхождение муравьев от одиночной (одинарной) семьи к федерации, мы можем наблюдать и параллельное развитие основного средства, обеспечивающего существование организационных структур как внутрисемейного, так и надсемейного ранга. Перенос особей, изначально связанный только с уходом за молодью в примитивных гнездах, а позднее - с переселением при смене гнезд, в дальнейшем вылился в мощное средство, без которого развитие социальности у муравьев трудно представить.
Сначала обмены открыли возможность возникновения в семье колонн. Затем, преодолев зачаровавший натуралистов отрицательный фототаксис, потоки муравьев-носильщиков вынесли молодь из гнезда и проложили путь к колонии. И наконец, интенсификация обменов позволила решить проблему «несовместимости» колоний и создания федерации.
В заключение данной главы следует отметить, что колониальность у муравьев изучается пока что на весьма ограниченном числе видов, относящихся в основном к различным подродам нескольких родов. Столь же ограничен круг поликалических видов, в той или иной мере изученных со структурных позиций. Степень изученности отдельных вопросов организации семьи, ее структуры, различна и не позволяет представить в настоящее время картины целиком даже у наиболее изученных в этом отношении муравьев. Отрывочные данные по структуре поселений других видов позволяют лишь предполагать степень сходства подобных явлений у других групп муравьев.